Acknowledgements
I extend my thanks to S. Kuzma for library assistance; M. Mayer and J. Harris for secretarial assistance; F. Gaynor for computer advice; and L. Roberts and Dr. K. Shapiro for editorial advice. I also thank C. Quinn for typing early versions of the manuscript, and the late E. Seiling and A. St. Laurent of United Action for Animals for suggesting ways to monitor and summarize animal research. I am especially grateful to the American, National, and New England Anti-Vivisection Societies, who commissioned this report, for the opportunity to conduct this study.
Forward
This is the second in a series of critiques of the strategy of animal models in psychological research.* The purpose of the present monograph is to provide a comprehensive view of a particular area of study—the investigations of primate maternal deprivation begun in the 1950s by Harry Harlow and recently given major support through the appointment of Stephen Suomi, a former coworker of Harlow’s, as director of a new National Institute of Mental Health primate facility.
The presentation of an overview of a discrete research area signals an advance in the contemporary reevaluation of science’s use of nonhuman animals as subjects. Since Singer’s broad philosophic critique (1975), the dialogue of charges and countercharges has ranged in targets from the entire field to the single study. In terms of sophistication of analysis, most recently the dialogue has moved away from selective field-wide cataloguing of either benefits to humans or costs to animals to citation analysis, which in this context means the documentation of the frequencies with which animal-based research is cited in the literature. For example, on the side of research benefits, Miller (1985; Coile and Miller, 1984) has surveyed a limited set of American Psychological Association journals to buttress his claim that much research involving animals is noninjurious and that it results in concrete applications. There is clear and compelling evidence that discounts both of these claims (J. Kelly, 1985; M. Giannelli, 1985). Unfortunately, to date the profession’s control over avenues of publication has prevented the dissemination of these rebuttals in the principal professional journals. Taken together, Kelly and Giannelli present a convincing case through data from citation analysis that Miller’s and other claims are unwarranted both with respect to degree of injury to animals and clinical applications for humans.
For this reason alone we welcome the use of the resources of the animal rights community in bringing before the public a more balanced understanding of psychology’s use of animal subjects.
In addition, the publication of a comprehensive analysis of the literature of maternal deprivation is a very fortunate choice for reasons Stephens indicates directly in his introductory chapter. At this point in the current dialogue we must complement the philosophic discussion, the general survey and the singular instance of alleged abuse with evaluations at the level of areas of research. While large, the number of studies in the area of maternal deprivation employing primates lends itself to such an evaluation: although ongoing for over thirty years, the lines of its development are readily grasped; it has been the object of criticism on ethical grounds by the animal rights community but at the same time almost every litany of “benefits” includes reference to it; it is an exemplar case of the animal model strategy of psychological investigation, a strategy that itself has become a significant object of debate.
Stephens’ effort excels in its clarity and its dispassionate tone. Much of the text concretely portrays this set of over 250 studies in an organized and straightforward manner that makes the research program and its impact on the animal subjects available to expert and layperson alike. While often simply allowing the record of the literature, including critical discussion by the researchers, to speak for itself, Stephens also provides his own critique. It is incisive and yet stays close to the facts.
We learn that the effects of this research have been profoundly injurious to over 7,000 animals. Despite this “cost,” direct applications from the research are virtually nonexistent. Stephens is able to trace only one application, which he then finds is an effective intervention for reasons other than those suggested by the research. We learn further that since its inception maternal deprivation research has largely been the offspring of one “family” of researchers, predominantly students of Harlow, trained by him at the University of Wisconsin. One implication of this surprisingly inbred genealogy is that this particular research program is not the inevitable process of impersonal institutional forces that have a momentum of their own, but rather is the result of the momentous personal conviction of a small group of individuals. In this context, Stephens argues that the preponderance of the studies represent no more than “parametric tinkering” and unnecessary elaborations on Harlow’s original work.
But what of the theoretical advances and the conceptual clarifications provided by Harlow’s paradigmatic experimental design—disrupting the primate mother/infant relation? Surely these begin to justify this research. As a clinician, I am not surprised at Stephens’ finding of few direct applications from this research to the treatment of depressed, isolated, or abused individuals. However, as Midgely (1981) has pointed out with respect to various psychological research areas, readers approaching this literature, both psychologists and others, assume that at least there is much gain here theoretically, that both major theoretical disputes and heretofore unnoticed conceptual distinctions were settled or crystallized through it. “I had assumed that a deeper structure of thought must be present” (p. 332). But her examination of, for example, Harlow’s studies of “chambering” as a model for the production of human depression reveals them to be based on vague intuitions of similarities and to fail to provide any sharpening of our thinking about depression. In his comprehensive analysis Stephens confirms this. With the exception of the critique of the “primary drive” theory of motivation, a critique provided by numerous other research findings by the 1950s, there has been little theoretical payoff from the maternal deprivation studies. Psychoanalytic theory and clinical studies, the work of Spitz and Bowlby, not to speak of common sense, all point to the pernicious effects of early deprivation and separation of child from parent and the positive effects of enriched contacts, including affectionate contact. Beyond this, the many particular variables examined in this research pro-gram have yielded very little information that is conceptually clarifying. Many of the particular findings, beyond the primary theme of the gross effects of deprivation, are not generalizable from rhesus to pigtail monkeys let alone from the former to humans.
None of the early hopes for discovering a useful animal model for depression or for maternal isolation have been realized. That they have not is not an indictment of the ingenuity of these researchers. The reason for the failure runs deeper. The strategy of developing animal models for phenomena, which are essentially a function of cultural arrangement and of linguistic mediation, is a poor one. This is not to say that nonhuman animals do not suffer from depression, anxiety, apprehensiveness about separation and the like. It is only to assert that the constitutive and determinative features of human depression, its forms and occasions and, undoubtedly, therefore, the keys to its treatment are linguistically and culturally embedded. It is not a question of “further research,” of more carefully teasing out confounding variables, or of replacing the blow of the sledgehammer with a few gentle taps (to use maternal deprivation researcher Mason’s critical image of animal models). To understand depression in our particular contemporary culture, we cannot simply tap, gently or otherwise, our nonhuman primate “resources”—not in any case and certainly not when those animals are bereft of those natural settings which they require to be themselves and therefore to show us their own complex natures.
Yet we cannot deny that Harlow and family have had an impact on psychology and even on our contemporary culture. In retrospect the primary impact of their work is the provision of graphic metaphors for the phenomena under investigation. Stephens asserts that, “many of these experiments were little more than sensational illustrations of what had already been established in humans” (p. 58). Considering psychology as a natural science, this is not a function that is justifiable. However, taking psychology as a peculiar institution within our culture, these studies have given us a set of metaphors: radical isolation as an infant primate reduced to a whimpering ball of fur, utterly and irreparably destroyed; rejection as in an unforgiving spiny-porcupine monster. These images are part of our culture now, part of us, and perhaps, even part of what we imagine when we are feeling isolated or depressed. We did not so much gain in understanding from these studies – for the metaphors have no independent exploratory value – as were we changed by them. On ethical grounds it is clear that we cannot justify the time, money, and gross exploitation of animals for “benefits” such as these.
Prospectively, I believe that the chief contribution of these unthoughtful sledgehammer attempts to solve complex human problems will be in the areas of ethics — for they will have demonstrated to us the cruel excesses of our use of and disrespect for other sentient and intelligent beings. Michael Foucault’s description of the administration of justice in eighteenth century France provides a historical parallel (1979). At that time the trial of the accused involved undergoing an ordeal, a systematized suffering. It was believed that only through such a “trial,” in the original sense of that term, and through the confession exacted in the process could the truth be established. In this primitive form of justice the magistrate did not intend cruelty, yet, in part, the callousness of this judicial system led to the upheavals of the French Revolution. Aspects of present-day American psychology, notably exemplified by the animal model investigations of psychopathology under study in this monograph, also involve unintended or at least incidental suffering in the service of truth seeking. In the not too distant future, we will have the same repugnance at this contemporary “primitive” science with its experimental manipulations in the name of truth as we do now at Foucault’s accounts of justice in eighteenth century France.
It is a common saying that nothing is as American as motherhood and apple pie—as innocent, as fresh, as life-sustaining, as good. Yet through these studies of motherhood we are taken to the lower reaches of a medieval Dantesque inferno. As the present monograph objectively and effectively demonstrates, this is the case because what shows through in this research is not the subject matter of the investigation but its method—a primitive form of truth seeking through suffering. Ultimately, what we gain from this extended investigation is not an understanding of the “nature of love” (Harlow’s term) nor even cultural metaphors for love or the effects of its absence. In the final analysis, we are given an image of a science, a science that has no recourse but to reform itself.
Kenneth J. Shapiro, Ph.D., A.B.P.P.
Bates College
Lewiston, ME 02440
*The first is Brandon Kuker-Reines, Psychology Experiments on Animals: A Critique of Animal Models of Human Psychopathology (1982).
References
Coile, D. and Miller, N. (1984). How Radical Animal Activists try to Mislead Humane People. American Psychologist, 39, 700-701.
Foucault, M. (1979). Discipline and Punishment: The Birth of the Prison. New York: Vintage.
Giannelli, M. (1985). Three Blind Mice, See how They Run: A Critique of Behavioral Research with Animals. In M.W. Fox & L.D. Mickley (eds), Advances in Animal Welfare Science 1985/86. Washington: HSUS.
Kelly, J. (1985). Personal communication.
Kuker-Reines, B. (1982). Psychology Experiments on Animals: A Critique of Animal Models of Human Psychopathology. Boston: NEAVS.
Midgely, M. (1981). Why Knowledge Matters. In D. Sparlinger (ed.), Animals in Research. New York: Wiley.
Miller, N. (1985). The Value of Behavioral Research on Animals. American Psychologist, 40, 423-440.
Introduction
During the last ten years, several critical evaluations of animal research have been written (Ryder 1975, 1983, Pratt 1976, 1980, Ruesch 1978, 1982, Diner 1979, Rowan 1984). These critiques have drawn attention to the plight of laboratory animals and have identified key issues in the controversy surrounding animal research. These key issues include: (1) the nature and extent of suffering inflicted on the animals, (2) the extent of unnecessary duplication of experiments, (3) the cost of the research to taxpayers, (4) the potential benefits to humans derived from the research, and (5) the potential alternatives to using animals in research.
Unfortunately, no field of animal research has been investigated to the extent needed to adequately address these issues. The books cited above were general surveys of animal research. Critiques of specific fields of animal research (Drewett & Kani 1981, Kuker-Reines 1982) addressed some of the issues mentioned above. Also, numerous reports provided examples of experiments that involved considerable animal suffering or that were of questionable value. However, as important as these reports are in exposing problems, they are not substitutes for comprehensive critiques. Compilations of experiments drawn from different fields are open to various countercharges. It could be claimed that the criticized experiments were taken out of context, and this distorted the rationale for the experimental design or the merit of the research. It could also be claimed that the experiments were not representative of studies in their field.
The lack of comprehensive critiques of specific fields of animal research is understandable because “research on research is unusually difficult and time consuming” (Comroe & Dripps 1976, p. 110).
This report is the first in a series of critiques designed to fill the need for comprehensive evaluations of particular fields of animal research. The report evaluates several areas of psychological research that fall under the heading of maternal deprivation. It addresses the issues of cost (both to the animals and to the taxpayers), benefits, alternatives and duplication. Virtually all maternal-deprivation experiments were funded by the federal government and were justified on the basis of their purported relevance to human mental disorders. Therefore, the conclusions of this report should be of interest not only to animal welfare advocates, but also to those concerned about human mental health and allocation of tax dollars.
Maternal deprivation research is noteworthy for several reasons. First, much of this research has been conducted on monkeys and apes. The experimental use of nonhuman primates, our closest relatives, raises particularly serious ethical issues. It is also highly controversial (e.g. Torrey 1984). Large demonstrations against primate research were held throughout the world on April 24th, 1983. Second, many (if not most) maternal deprivation experiments were conducted at Regional Primate Research Centers. These Centers were the targets of the 1983 demonstrations, and have also been criticized by scientists on bureaucratic grounds (see Rowan 1984, p. 125). Third, much of the maternal-deprivation research was directed by the late Harry Harlow, whose experiments and remarks have drawn scathing criticism from animal welfare advocates (e.g. Helton 1985). Rowan (1984, p. 141) noted that “most animal welfare literature that addresses psychological research on animals focuses on Harlow’s studies . . .” Finally, as a field of psychology, and not biomedicine, maternal deprivation research seems particularly difficult to justify on the basis of potential advances to human health (Hubbard 1985).
All of the experiments surveyed in this report involved taking infants from their mothers, sometimes at birth. Depending on the specifics of the experiments, the infants experienced emotional distress or psychological devastation; some died. Infants were sometimes subjected to procedures in addition to maternal separation. For example, monkeys at the Wisconsin Primate Center were separated from their mothers and incarcerated in isolation chambers or ‘wells of despair,’ or exposed to abusive artificial mothers. Some of the more fortunate infants at Wisconsin were left alone in tiny, barren cages.
For convenience, maternal deprivation experiments have been divided into three categories: affection, deprivation and separation. The affection category represents Harlow’s research on ‘the nature of love.’ In these experiments, monkey infants were deprived of their mothers at birth and reared with different types of artificial mothers. The aim of this research was to determine how infants became attached to their mothers.
The deprivation category, like the affection category, includes experiments in which infants were taken from their mothers at birth or shortly thereafter. In contrast to the affection studies, however, the emphasis here was on the effects of the deprivation experience. The deprived infants were usually reared alone, either in wire cages or isolation chambers. They were some-times reared with artificial mothers or other infants.
The final category of maternal deprivation, separation, is a heterogeneous collection of experiments that were once all considered to be animal models of depression. The majority of these experiments examined the consequences of separating an infant from its mother after an attachment bond had formed between them. Most of these separations were temporary. A similar type of separation procedure involved separating infants from other infants. Also included in the separation category, for convenience, are studies involving ‘wells of despair’ (vertical chambers) along with other procedures.
All three categories of maternal deprivation research, affection, deprivation and separation, are frequently discussed with reference to ‘attachment theory’ in psychology and psychiatry. Attachment refers to the emotional bond between individuals. In the context of maternal deprivation studies, the bond is usually that between infants and mothers.
In summary, the experiments surveyed in this report all involved removing mothers from their infants. The aim of these experiments was to determine the effects of this removal or to investigate other aspects of attachment behavior. Several related types of experiments are not included in this survey. These include (1) deprivation experiments not involving infants, and (2) experiments in which infants were removed from their mothers, but not for reasons other than those of interest here. For example, students of sexual behavior have often employed deprivation techniques (e.g. Shaw 1962), but their focus was sexual behavior, not deprivation.
Virtually all maternal-deprivation experiments were conducted since the early 1950s (Figure 1). Activity in the three fields of maternal deprivation research peaked in successive periods: affection experiments in the late 1950s, deprivation experiments in the mid-1960s, early 1970s and separation experiments in the 1970s. Affection experiments were much less common than deprivation and separation experiments (Figure 1). Most recent maternal deprivation studies are in the separation category; consequently, that category will be emphasized in this report.
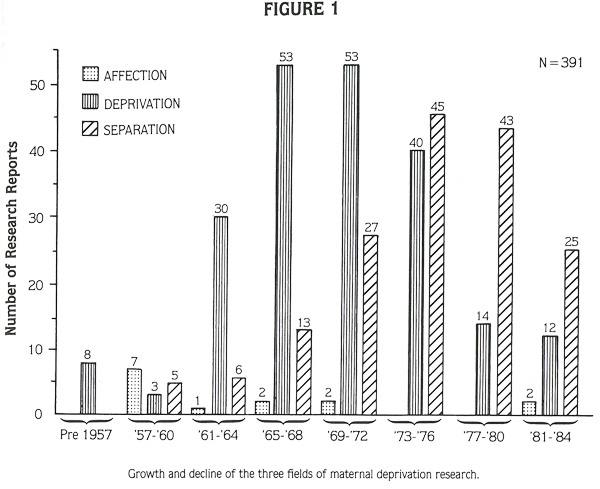
The next chapter (Chapter Two) provides background material on animal models in psychological research. Chapters Three through Five provide summaries and evaluations of each of the three categories of maternal deprivation research. The final chapter includes a general discussion and summary of the report’s findings.
Please cite as:
Stephens, Martin L. (1986) Maternal Deprivation Experiments in Psychology: A Critique of Animal Models.
Retrieved from http://aavs.org/maternal-deprivation-experiments-psychology.
ANIMAL MODELS
Why do psychologists use animal models of human clinical problems? One reason researchers turn to animals is that they are prohibited from conducting certain experiments on humans; the ethics of human research purportedly dictate that animals be used. The implicit assumption is that what can’t be done to humans, can be done to animals (e.g. McKinney 1974a, p. 530). Ethical considerations regarding animal research, when raised, are considered outweighed by potential human benefits.
Another rationale for conducting animal experiments is that certain experiments are more easily conducted on animals than on humans. For example, a prospective study of the effects of early experience on adult behavior may take twenty years to complete on humans, given their slow development, but only five years on monkeys.
Both the ethical and practical constraints on human research are apparent in Stephen Suomi’s justification for animal models:
… animal models can provide investigators with experimentally rigorous research tools with which ethically delicate and practically difficult questions about human psychopathology can be addressed (Suomi 1984a, p. 228).
One can ask why experiments that are prohibited or impractical on humans should be conducted on animals. The answer often involves the experimental psychologist’s reliance on highly manipulative, controlled experiments. Indeed, these psychologists sometimes convey a sense of disdain for clinical or naturalistic observations.
The use of animal models is not without its own difficulties, however. One obvious problem is that different species do not necessarily react the same way to a given experience. Therefore, conclusions about one species may not generalize to a closely related species, much less to humans. A second problem with many animal models is the artificiality of laboratory conditions, especially in research by experimental psychologists. These conditions yield results of questionable relevance to the natural characteristics of the animal model species, let alone those of humans.
The animal modelers themselves are increasingly recognizing shortcomings of animal models. Consider again the remarks of Suomi, who has conducted numerous maternal deprivation experiments and championed the use of animal models:
… in virtually no case is an animal model a perfect . . . replica of the human disorder under study Rather, it is usually a highly simplified, theoretically biased, and incompletely generalized version, expressed in members of a nonhuman species. An animal model is almost never the “real thing”; it is, instead, only a model of the real thing (1982, p. 250).
Therefore,
… the primary rationale for creating most animal models lies not so much in any obvious and impressive strengths of such models as it lies in the problems inherent in conducting research with humans as subjects (p. 250).
The experimental psychologists are therefore attempting to make the best of a bad situation. They can’t conduct their reductionist experiments on humans, yet they don’t have good substitutes for human subjects. This predicament is reflected in Harlow’s adage that “you’re crazy to use animal models and you’re crazy not to” (Kornetsky 1977, p. 498).
How can animal modelers deal with the problems inherent in using animal models? McKinney and Bunney (1969) provided a set of guidelines for modelers of human mental disorders. According to these authors, the animal model and the human disorder should have similar, if not identical, (1) causes, (2) symptoms, (3) biological mechanisms, and (4) cures. The more criteria satisfied by the model, the greater its scientific validity. Unfortunately, these criteria are often overlooked in maternal deprivation research.
-psychology/chapter-three/”>n style=”color: #808080;”>
Please cite as:
Stephens, Martin L. (1986) Maternal Deprivation Experiments in Psychology: A Critique of Animal Models.
Retrieved from http://aavs.org/maternal-deprivation-experiments-psychology.
EXPERIMENTS IN AFFECTION
Maternal deprivation experiments in the affection category were attempts to determine which maternal characteristics elicit an infant’s emotional attachment to its mother. All but two of these experiments were conducted on rhesus monkeys at the University of Wisconsin and Wisconsin Primate Center. Harlow directed this research, labeling the topic “the nature of love.” His experiments involved depriving rhesus monkey infants of their mothers at birth and rearing them with inanimate surrogate mothers.
Overview of Affection Experiments
Harlow began his affection research after depriving newborn monkeys of their mothers and noting that they became psychologically attached to cloth diapers placed in their cages. The infants’ attachment to cloth diapers gave Harlow the idea of using cloth surrogate mothers to demonstrate which maternal attributes elicited filial affection.
Harlow’s initial study (Harlow 1958, 1959a, 1962a, 1962b, Harlow & Zimmerman 1959) attempted to determine which of two factors was more important in eliciting filial affection. One factor was the physical contact associated with clinging to the mother. Harlow referred to this as “contact comfort.” The other factor was the breast and activities associated with nursing.
Harlow’s observations of infants becoming attached to cloth objects led him to suspect that contact comfort was an important factor in filial bonding. He pitted contact comfort against nursing as a challenge to prevailing views on the importance of nursing in filial attachment. For example, the prevailing view of psychologists was that human infants learned to love their mothers by associating the mother with the satisfaction gained from suckling. In the terminology of learning theory, mother’s milk was the primary reinforcer that satisfied the infant’s hunger, and mothers themselves were the secondary reinforcer. Accordingly, this view of filial bonding was known as the Reinforcement Theory. It was also known as the Drive Reduction Theory, a reference to the infant’s hunger as a biological drive.
In Harlow’s initial study he deprived rhesus monkey infants of their mothers and reared them with both a cloth surrogate mother and a wire surrogate mother (Photo 1). A nursing bottle was attached to one surrogate or the other, but not both. The comparison of greatest interest was the cloth surrogate without the bottle versus the wire surrogate with the bottle. This pitted contact comfort against nursing. A control group, referred to as “orphans” (Harlow 1959a), was denied physical contract with any surrogate mothers or other monkeys for about eight months from birth.
Several lines of evidence suggested that contact comfort was more important than nursing in eliciting filial attachment. For example, infants spent more time with the cloth surrogates (CS) than with the wire surrogates (WS). Other measures of the infants’ greater attachment to CSs clearly involved distressing procedures. In the “home cage fear test,” an opaque screen in the infants’ cage was suddenly lifted, exhibiting a “mechanical monster” (Harlow & Harlow 1965). The monster moved towards the infant while making loud noises. This produced “abject terror” in infants, who frantically ran to their CSs (Harlow & Zimmerman 1959).

Rhesus monkey infant housed with surrogate mothers made of wire and cloth. From Harlow (1966) with permission of Harper & Row.
Another distressing test involved placing infants in a large, unfamiliar room containing several objects. In the presence of their CSs, infants would rush to the surrogates in fear. If the CSs were absent, sometimes the infants would run to where the CSs customarily were and then run
from object to object, screaming and crying all the while. Continuous, frantic clutching of their bodies was very common . . . (Harlow 1958 p. 680).
Also, the infants would
rush across the room and throw themselves facedown on the floor, clutching their heads and bodies and screaming their distress (Harlow 1959a, p. 72).

Rhesus monkey infant after being placed in a strange room without its cloth surrogate mother. From H. F. Harlow, 1958, “The Nature of Love,” Am. Psychol. 13:680. Copyright (1958) by the American Psychological Association. Reprinted by permission of the publisher.
These infants eventually were separated from their CSs and later tested to see if they had retained their affection for the surrogates. The tests administered during early infancy were repeated and yielded similar results. One variation of the test in the unfamiliar room involved shielding the CSs, when present, by a plexiglass box. Hence, the youngsters could not reach them. The infants crashed into the plexiglass in frantic efforts to reach their surrogates. A more frightening variation of the fear test was also devised: a plexiglass panel forced infants to approach the mechanical monster in order to reach their cloth mothers.
The “orphans” initially deprived of real and surrogate mothers were later housed with CSs and WSs, neither of which supplied milk. The orphans’ initial reaction to the strange surrogates was one of “extreme disturbance.” “All infants screamed violently and made repeated attempts to escape the cage whenever the door was opened” (Harlow 1958, p. 683). However, the orphans eventually became attached to the CSs.
According to Harlow (1959a), the results of his initial experiments indicated the “surprising importance of contact comfort as a prime requisite in the formation of an infant’s love for its mother” and the “unimportant role of the breast and act of nursing” (p. 74). These results were considered a fatal blow to the Drive Reduction Theory, which stressed the importance of nursing.
Harlow’s follow-up experiments were less consequential. These involved rearing infants with (1) a CS or a nursing WS, but not both (Harlow & Zimmerman 1959), (2) a CS and a nursing CS (Harlow 1960), (3) a rocking and a non-rocking surrogate, (4) a “clingable” and a non-clingable surrogate (Harlow 1959a, Harlow 1962), and (5) surrogates covered by cotton, rayon, vinyl or sandpaper (Furchner & Harlow 1969).
Harlow and Suomi (1970) conducted the most recent affection experiments at Wisconsin. Much was made of the new, simplified design for surrogate mothers a terry cloth torso with an optional head. The researchers again assessed the importance of several maternal variables in the development of filial affection.
One such variable was nursing. Harlow and Suomi admitted that one of Harlow’s major conclusions that activities associated with the breast were of no importance—was “undoubtedly incorrect.” The new experiments showed that infants preferred a nursing surrogate over a non-nursing one, though the preference waned over time. Harlow and Suomi did not note that Harlow reported the same finding in 1965 (Harlow 1965).
Another experiment reported by Harlow and Suomi showed that body warmth is an important affectional variable. One of these experiments involved surrogate mothers whose bodies could be warmed and chilled experimentally. An infant huddled in a corner and cried “piteously” when its surrogate was chilled. Harlow and Suomi, “feeling somewhat guilty,” restored the warmth, but later resumed alternating warm and cold temperatures. A second infant was subjected to a modified procedure and was so devastated that it did not even cling to the surrogate when the latter was warm.
One affection study was conducted on dogs (Igel & Calvin 1960). “Mongrel” puppies were separated from their mothers at birth and reared with a CS, a WS, or both. The amount of time spent on each surrogate was the sole criterion for the strength of attachment.
As in the Wisconsin studies, ‘contact comfort’ was found to be a highly important variable, but in the present study, contrary to the Wisconsin finding, lactation was found to have a very marked effect for the subjects with cloth mothers (Igel & Calvin 1960, p. 305).
Hence, the researchers believed that their results revived the notion that nursing is a factor in filial bonding. Harlow reached a similar conclusion after this dog experiment was published (Harlow & Suomi 1970, see above).
Discussion
The Nature and Extent of Suffering
Over 62 animals were involved in the affection experiments (Reference Note 1, p. 11). Most of these animals were the monkeys in Harlow’s experiments. Defenders of Harlow’s experiments might claim that these monkeys were not harmed. However, Harlow’s experiments were quite stressful and psychologically damaging. Virtually all of the infants were, in Harlow’s own words, “terrorized” in various tests.
Much of the trauma in these experiments could have been avoided. For example, one can argue that the tests used to demonstrate the superiority of cloth surrogates over wire surrogates were unnecessarily stressful and excessive. Simply the amount of time the infants spent on each surrogate would have been a sufficient measure of the cloth surrogate’s superiority (as in the dog study). Even if the supplementary tests were considered essential, they could have been modified to be less stressful but equally convincing.
The full extent of the damage inflicted in these experiments was not revealed in Harlow’s original reports. The extensive disturbance of surrogate-reared infants was realized only when these infants grew up. Rhesus infants reared with inanimate surrogate mothers became as psychologically disturbed as infants reared alone (see Ch. 4). One manifestation of their abnormality was “chewing or tearing at [their own limbs] with the teeth to the point of injury” (Harlow & Harlow 1982a, p. 217). Ironically, Harlow initially boasted that his surrogate mothers were “mother machines” that were superior to real mothers.
Some of Harlow’s infants not only were deprived of their real mothers, but also prematurely separated from their surrogate mothers. Such separations can induce anxiety and grief.
Finally, the suffering of monkey mothers should also be considered. These mothers no sooner bore their offspring than the latter were taken from them. The removal procedure involved a struggle between the mothers and the researchers. In the aftermath of this struggle, the mothers undoubtedly grieved over the loss of their infants.
Unnecessary Duplication
Do affection studies contain unnecessary duplication? This issue is separate from the question of whether or not this line of research should have been conducted at all. Several reports of affection research published by Harlow were largely different versions of the same manuscript, making the research appear more duplicative than it actually was. However, some of Harlow’s latest experiments (Harlow & Suomi 1970) duplicated earlier research.
Financial Cost
The Wisconsin affection experiments cost approximately $2.4 million to conduct (Reference Note 2, p. 12). (No funding information is available on two studies not conducted at Wisconsin). Ninety six percent of this amount was supplied by the National Institutes of Health. This means that American taxpayers paid for virtually all of Harlow’s affection research. Lesser amounts were supplied by the Ford Foundation ($100,000) and the University of Wisconsin (exact amount unknown).
Benefits to Humans
Harlow’s affection experiments brought him scientific acclaim. The most widely cited finding was that rhesus monkey infants preferred a cloth mother to a wire mother, even when the latter provided milk. This finding discredited the Drive Reduction Theory of how humans develop an attachment bond to their mother.
The idea that infants’ emotional ties to the mother . . . [was] based on the reduction of biological drives dominated American theories of infancy from World War I until the early 1960’s. Because the feeding situation was considered so important, child development experts and parents devoted a great deal of attention to whether a child was breast-fed or bottle-fed, whether fed on schedule or demand, [etc…]. Although these important questions were investigated extensively by scientists, no consistent relations between feeding patterns and the child’s subsequent social and emotional development were discovered. . . . But a more fatal blow to theories emphasizing biological drive reduction was dealt by Harry Harlow and his colleagues (Mussen et al. 1984, p. 170).
Hence, Harlow’s affection experiments on rhesus monkeys were considered to have advanced the understanding of human infancy. This theoretical shift undoubtedly influenced later research on infant development in humans. However, there is no apparent evidence that Harlow’s affection experiments have had any impact on clinical practice, either directly or indirectly.
Harlow and Contact Comfort
The concept of contact comfort is most closely associated with Harlow, and rightly so. However, Harlow’s contribution in this area should not be exaggerated.
Harlow was not the first to recognize the phenomenon of contact comfort, nor the first to suggest it was important in bonding human infants to their mothers. Psychoanalysts did not use the term ‘contact comfort,’ but they clearly referred to the same phenomenon. Indeed, some psychoanalysts postulated a clinging drive that was capable of bonding infants to mothers (Bowlby 1958). This view, also shared by ethologists, was the major alternative to the Drive Reduction Theory in the period before Harlow began his affection experiments.
Contact comfort in infant monkeys and apes was recognized decades before Harlow’s work (Wallace, cited in Mason 1968; Foley 1934, 1935). Foley related this “contact clinging” and its emotional consequences to the rhesus monkey’s semi-arboreal existence. Harlow seemed reluctant to acknowledge the obvious connection between infant clinging and arboreality. Perhaps Harlow thought that this connection revealed that rhesus monkeys may not be a good model of filial attachment in humans. Harlow’s contribution to these earlier studies was to provide evidence that the importance of contact comfort in infant primates was counter to prevailing views of filial attachment.
This evidence was recently criticized, on methodological grounds, by L. Ainsworth, who conducted his own affection study (Ainsworth & Baker 1982). His myth-shattering conclusion was that:
Contrary to general belief, published reports of the original infant/surrogate work contain few data to support the idea of a need for contact comfort . . . (Ainsworth 1984, p. 943).
Harlow’s Conclusions and Their Implications for Humans
Perhaps Harlow’s most surprising and influential conclusion was the “unimportant role of the breast and act of nursing” in the formation of an infant’s love for its mother (Harlow 1959a, p. 74). Yet Harlow himself later admitted he was “undoubtedly wrong” in dismissing the importance of nursing (Harlow & Suomi 1970). Harlow came to recognize nursing as well as contact comfort as important variables binding rhesus monkey infants to their mothers. Bowlby had reached a similar conclusion regarding human infants in 1958, although he admittedly lacked experimental data.
Harlow was initially led astray on the importance of nursing probably because his nursing surrogate mother was made of wire. The wire may have so frustrated the pervasive clinging responses of newborn primates that the importance of nursing could not be manifested (Gerwitz, in Harlow 1965).
Even if clinging is more important that suckling in filial attachment in rhesus monkeys, the same is not necessarily true for humans. Mason, a former student of Harlow, cast doubt on the general relevance of monkey affection data for humans.
No human infant, of course, is able to cling, find the nipple, and to nurse without extensive maternal assistance. The same infantile responses that make the difference between life and death for the rhesus monkey [e.g. clinging] are often viewed in the human infant as mere behavioral curios, without obvious adaptive value (Mason 1968, p. 98).
Mason (1968) concluded that animal models of human behavior work best for well-circumscribed processes and not, for example, “the origins of love.” This candid statement is a clear reference to Harlow’s affection research, which Harlow did consider to be an animal model of love (Harlow, in Tavris 1973). Mason presumably would agree with psychoanalyst Dallas Pratt (1980), who wryly noted the following:
It has been pointed out that contact comfort is more important in monkeys than in man, and anyone who has watched an infant monkey clinging tightly to its mother as the latter swings through the trees can easily understand why (p. 61).
To summarize, Harlow’s animal model of filial affection overestimated the importance of contact comfort and underestimated the importance of nursing. Hence, any direct applications of Harlow’s conclusions to humans may have led human research astray.
Alternatives
It has already been suggested that Harlow could have modified his procedures to reduce the stress to which he subjected his monkeys. Could he have avoided using animals altogether? While no one would suggest depriving human infants of their mothers to conduct affection research, certain types of human studies can be conducted. For example, mothers could provide variable levels of contact comfort or nursing while keeping the other factor constant. Thus, all infants would thereby receive contact comfort and nursing, but some would receive enhanced levels. The strength of the infants’ developing attachment to their mother could then be monitored. Psychologist Mary Ainsworth has conducted studies along these lines. For example, on the basis of one of Ainsworth’s recent studies, Mussen et al (1984, p. 170) concluded that “the strength of a child’s attachment to either parent is not related in any simple way to the frequency with which that parent feeds [the child].”
In the 1950s, when Harlow’s first affection experiments were conducted, infants in many orphanages lived in severely deprived environments (Bowlby 1952). This unfortunate situation could have provided opportunities for positive, albeit experimental, intervention by caretakers and researchers into the nature of filial attachment.
Summary
Affection experiments involved rearing infant monkeys and dogs with artificial mothers having different characteristics. The aim of these experiments was to determine which maternal characteristics underlie the infant’s attachment to its mother.
Most of the 100 or so animals involved in these experiments were infant monkeys. They were distressed in various tests and psychologically damaged by maternal deprivation. The nine studies conducted by Harlow cost a total of about $2.5 million, supplied primarily by the federal government. The studies had an impact on child development theory, but no direct impact on human welfare. Some alternatives to using animals in this research are cited.
Reference Notes
1. Throughout this report, numbers of animals refer only to those animals subjected to deprivation experiences. Animals not subjected to these experiences, such as animals in some control groups, were not included in the totals. Furthermore, an effort was made to avoid recounting the same animals mentioned in two or more reports.
2. Determining the exact cost of particular studies is difficult. Individual studies are parts of larger research projects, and these projects are the recipients of research grants. Funding data cited throughout this report are therefore approximate. The cost of some studies undoubtedly has been overestimated, but this should be offset by studies whose cost has been underestimated, or by studies for which funding data are lacking.
Please cite as:
Stephens, Martin L. (1986) Maternal Deprivation Experiments in Psychology: A Critique of Animal Models.
Retrieved from http://aavs.org/maternal-deprivation-experiments-psychology.
Experiments in Deprivation
The severe psychological problems that developed in Harlow’s monkeys led him to consider the effects of maternal deprivation. Hence, Harlow’s experiments in the affection category led to his experiments in the deprivation category. These experiments concern the effects of social deprivation on infant development. In virtually all of these experiments, infants were not only deprived of their mothers, but also denied contact with any other individuals of their species. They were kept alone in bare, wire cages or in isolation chambers.
Several rationales for deprivation experiments have been offered by those conducting this line of research. These experiments have been considered to be animal models of social isolation, mental illness, or even normal development. Some have been considered to be relevant to child abuse. In addition, some were attempts to assess the importance of early life experiences to behavioral development. Rearing an animal in a depriving environment was a means of nearly obliterating its life experiences. Hence, any behavioral abnormalities that developed were attributed to deficient experience, not deficient genetic endowment.
The following section is a summary of these deprivation experiments. It should be emphasized that our concern here is not with deprivation experiments in general, but with those that deprive infants of their mothers. Deprivation experiments on juveniles and adults are beyond the scope of this report.
Overview of Deprivation Experiments
Deprivation experiments have been conducted primarily on nonhuman primates and dogs, and only these studies are considered here. Some deprivation experiments have been conducted on farm animals and rodents but these studies are all but ignored by researchers studying dogs and primates. The studies of farm animals are outdated and those on rodents mostly involved animals older than infants. For reviews of the experiments on farm animals and rodents, see Moore (1968), Porter (1974) and Thoman and Arnold (1968).
Experiments on Dogs
Several deprivation experiments were conducted at McGill University during the 1950s. These experiments involved rearing terriers in isolation chambers for several months (Photo 3). Isolation began when the puppies were separated from their mothers at four weeks of age. The emphasis of these experiments was on the consequences of early experience for behavioral development.
The results indicated that isolation-reared dogs had bizarre “whirling fits” (Thompson et a/. 1956) and aberrant, highly excited responses to innocuous objects (Melzack 1954). Hebb, a McGill researcher, wryly described these animals as “permanent screwballs” (Hebb 1958, p. 109). The isolates were poor at problem-solving, “a result in agreement with previous animal experimentation” (Thompson & Heron 1954, p. 31). They were also inferior to nonisolated dogs in social behavior, even when reared with a companion in their isolation chambers (Melzack & Thompson 1956).
 Photo 3
Photo 3
Isolation chambers for dogs at McGill University. From R. Melzack 1965, “Effects of early experience on behavior,” in Psycho-pathology of Perception (P H. Hoch & J. Zubin, eds.), Grune & Stratton, Inc. Reproduced by permission of author and publisher.
Melzack continued these experiments when he moved from McGill to the Massachusetts Institute of Technology (Melzack 1962, 1965, Melzack & Burns 1963). Isolation-reared beagles, like the McGill terriers, were hyperactive when removed from their chambers. They displayed abnormal responses to visual, auditory and olfactory stimulation, and seemed unperturbed after inadvertently crashing into things. Melzack attributed their difficulties to an inability to filter out irrelevant sensory information. This led to hyperexcitement, which in turn, interfered with testing performance. Preliminary findings from physiological experiments tended to support this view.
Several deprivation experiments involving dogs were conducted at the Jackson Laboratories, a supplier of animals to research institutions. Puppies were reared in total isolation for three to five months, beginning at three or four weeks of age. As in the McGill and MIT experiments, the puppies were hyperexcited on emerging from their chambers. Fuller and Clark (1966a) characterized this behavior as a “panic response.”
…its behavioral manifestations can vary from cowering withdrawal, rigid immobility in unusual postures, through stereotyped approach-avoidance, and violent tantrums with loss of [bladder] control (p. 256).
Fuller and Clark’s experiment suggested that the effects of isolation resulted from the trauma of emerging from isolation, rather than from deprivation during the isolation period. They called their explanation the “emergence trauma hypothesis,” and contrasted it with the “critical period hypothesis.” The latter, when applied to isolation experiments, states that the ill effects of isolation result from a lack of normal experience during a “critical period” in development.
It is important to note that two breeds were tested and found to differ in the severity of their response to isolation. Hence results from one breed do not necessarily generalize to another breed, let alone to another species. The same conclusion was supported by another experiment (Fuller & Clark 1968).
Fuller and Clark (1966b) found that various forms of stimulation during isolation-rearing did not prevent abnormal social and exploratory behavior from developing. The authors interpreted these results as further support for the emergence trauma hypothesis.
Later experiments, reviewed in Fuller (1967), showed that less severe deprivations can have less severe effects. Fuller also reviewed experiments on the effects of isolation on problem solving. The results admittedly were inconsistent.
Several isolation experiments on dogs were also conducted at Galesburg State Research Hospital by Fox and coworkers (Fox & Stelzner 1966a). Isolates were caged individually and kept in a darkened room for one week, beginning at four weeks of age. Thus the pups experienced sensory as well as social isolation.
One week of isolation produced less drastic and persistent abnormalities than the long-term isolation in the McGill, MIT and Jackson experiments (Fox & Stelzner 1966b). For example, the isolates did not exhibit the “mass-fear response” (extreme avoidance and fearfulness) seen in the long-term isolates. The researchers attributed the differences between the long- and short-term isolates to the timing and the duration of isolation.
One week of isolation was sufficient to produce intense behavioral arousal on emergence. This arousal was associated with changes in brain physiology (Fox 1967). One group of dogs in this study had electrodes chronically implanted in their brains. The results lent support to the interpretation of the McGill, MIT and Jackson studies; namely, that dogs reared in isolation are unable to readily adapt to the sensory stimulation of normal environments.
Another experiment attempted to correlate behavioral and neurochemical effects of isolation (Agrawal et al. 1967). After isolation, the puppies were decapitated and their brain chemistry was analyzed. Several changes had occurred in the concentrations of free amino acids. The interpretation of some of these neurochemical changes was unclear. As the researchers noted, some may have resulted from the darkened isolation room, rather than the social isolation itself.
Experiments on Primates
University of Wisconsin and Wisconsin Regional Primate Research Center
Deprivation experiments on primates began in the 1930s (Foley 1934, 1935) but did not become popular until the 1960s. Most deprivation experiments on primates were conducted at the University of Wisconsin, under the direction of Harry Harlow. All of the Wisconsin deprivation experiments involved rhesus monkeys. These experiments were viewed as animal models of human development— both normal and abnormal. The model of normal development was viewed as a means of assessing the importance of early social experiences. Most of the attention, however, was on abnormal development, with the deprivation experiments being considered as models of maternal deprivation, social isolation and mental illness.
The Wisconsin researchers distinguished two types of isolation conditions. ‘Partial isolation’ involves rearing infants alone in bare wire cages. The infants can see and hear other monkeys, but not touch them (Photo 4). ‘Total isolation’ involves rearing infants in isolation chambers that preclude all forms of interaction between monkeys (Photo 5). The partial isolation experiments are summarized first, followed by the total isolation experiments, and then various other kinds of deprivation experiments are considered.
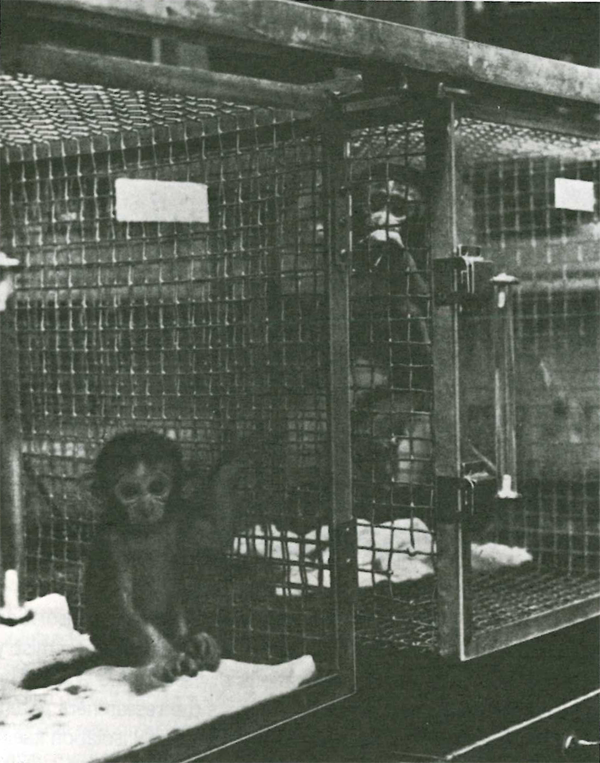 Photo 4
Photo 4
Rhesus monkey infants being reared in ‘partial isolation.’ Each infant is housed alone in a wire cage (the front panel of the cage is plexiglass). From Harlow and Harlow (1966) with permission.
Partial Isolation
Mason, who at the time was Harlow’s student, conducted the first Wisconsin experiments on the effects of partial isolation (Mason 1960, 1961a, 1961b, 1963a, 1963b, Mason & Green 1962). Rhesus monkey infants were subjected to partial isolation for their first 28-29 months. They were then tested in a variety of situations. A control group consisted of monkeys captured in India, shipped to a zoo, and later taken into the laboratory.
 Photo 5
Photo 5
Rhesus monkey infant in an isolation chamber. The photograph was taken when the chamber door was raised for the first time after half a year of ‘total isolation.’ Reproduced from Harlow (1964) with permission; copyright by the Williams & Williams Co., Baltimore.
The isolates were socially abnormal. They responded poorly to social cues, and their grooming, sexual, and aggressive behavior were impaired. They were also more disturbed than wild-born monkeys when placed in novel situations. Mason concluded on the basis of a variety of tests that “orderly and harmonious social relations in rhesus monkeys are dependent upon previous socialization experience” (Mason 1961a, p. 290).
Harlow and Harlow (1962b) described the abnormalities of a much larger group of monkeys reared in partial isolation. Fifty-six rhesus infants were available for study during the early 1960s because husbandry conditions that were routine at the time called for partial isolation. The unanticipated abnormalities of these isolates included blank staring and stereotyped, repetitive circling of cages. These infants often had compulsive habits, including self-mutilation. Harlow and Harlow (1971) described the self-mutilation of limbs:
Ordinarily [self-biting does] not break the skin, but under conditions of stress or threat they may literally tear their limbs to pieces. In a considerable number of instances, injury has been so severe that the animals had to be sacrificed (p. 210).
Surprisingly, partial isolates were nearly as proficient on learning tasks as were control monkeys (Gluck et al. 1973).
Isolates that were left in their cages did not outgrow their affliction (Cross & Harlow 1965; Suomi et al. 1971). They remained withdrawn and self-abusive although some developmental changes occurred (e.g. Arling et al. 1969). The control group in the Suomi et al. study had been born in India, captured and brought into the laboratory before their second birthday, and subjected to solitary confinement for 9 to 15 years.
Total Isolation
In the earliest series of Wisconsin experiments on the consequences of total isolation, newborn rhesus monkeys were confined for 3, 6 or 24 months (Harlow & Harlow 1962b), as well as for 18 months (Mason & Sponholz 1963). Three months of confinement had no noticeable effect on seven of eight infants. However, longer periods of confinement had increasingly severe effects. The 24-month old infants “were totally destroyed . . . ” (Harlow 1964, p. 159).
These initial experiments demonstrated that total isolation left monkeys extremely fearful (Photo 6). Indeed, “A stimulus such as a tennis ball . . . is apt to elicit intense terror from an isolate monkey,” even one reared in partial isolation (Suomi & Harlow 1976, p. 20).
 Photo 6
Photo 6
Rhesus monkey infant in isolation chamber. The photograph was taken after the chamber door was raised. Note the self-clutching, a characteristic of rhesus monkeys reared in isolation. From Harlow and Harlow (1965) with permission.
According to Harlow et al. (1965), newborns were also “sentenced” to 3, 6, or 12 months of total isolation. A fourth group was subjected to 6 months of isolation beginning at 6 months of age (“6-month late isolates”). Following the clinical literature, Harlow referred to the condition of the 6-month late isolates as ‘deprivation’ and that of the other isolates as ‘privation.’ No mention was made of the earlier Wisconsin experiments that sentenced newborns to 3, 6, 18, and 24 months of total isolation. Apparently, these experiments were regarded as preliminary.
The goal of the newer research was a better understanding of the effects of “human social isolation,” which was said to occur in orphanages, foster homes, and abnormal private homes.
The results of these experiments, which were reported by Harlow et al. (1964), Harlow et al. (1965), and Griffin and Harlow (1966) were as follows. When released from their isolation chambers, the infants initially suffered “emotional shock.” The three-month isolates were especially traumatized at release, showing “extreme withdrawal” and “severe depression.” This reaction was so severe that one [monkey] died of starvation because it refused to eat .. . (Griffin & Harlow 1966, p. 534). Another three-month isolate would have died of this “emotional anorexia” had it not been force-fed.
Total isolation had dramatic, debilitating effects on the survivors (Photo 7). The severity and persistence of these effects were related to the duration and timing of isolation. The effects were eventually reversed in the three-month isolates. The six months of early isolation “severely impaired” the potential for socialization, whereas “12 months of isolation almost obliterated the animals socially . . . ” (Harlow et al. 1965, p. 94). These asocial animals were severely disturbed. “Testing . . . had to be discontinued . . . because the control animals were mauling and abusing the helpless isolates to the point that they could not have survived continuing interaction” (Harlow & Harlow 1969, p. 18).
 Photo 7
Photo 7
A rhesus monkey infant reared in an isolation chamber. Reproduced from Harlow (1974) with permission.[/caption]
The 6-month late isolates showed neither the severe short-term reaction of the 3-month isolates nor the severe long-term reaction of the 6-month early isolates and the 12-month isolates.
Learning tests revealed little evidence of intellectual impairment in any group; extensive follow-up testing supported this conclusion (Harlow et al. 1971a).
These monkeys were 12 to 20 months old when tested initially. The same isolates were tested again at 28 to 44 months and showed few signs of improvement (Mitchell et al. 1966). They were fearful and aggressive and showed little or no interest in sex or play. Their acts of aggression were often suicidal attacks against huge adult males or brutal beatings of juveniles . ” (p. 578). Throughout testing, the 12-month isolate showed “practically no positive social behavior” (p. 568). The isolates were also characterized by “infantile disturbance” and bizarre ritualistic movements.
Mitchell (1968a) tested these isolates again when they were 42 to 54 months. There were some signs of improvement, though the isolates remained socially inept. Fear was increasingly replaced by hostility.
Mitchell and Clark subjected rhesus infants to six months of total isolation beginning at three months of age, instead of at birth or at six months of age as in the earlier Wisconsin studies. This study also differed in other ways from most earlier Wisconsin studies. One difference was that the isolates were reared with cloth surrogate mothers. Another difference was that the isolates were preadapted to the post-isolation test apparatus. The latter procedure was an attempt to reduce the trauma associated with emerging from isolation chambers. This part of the experiment was a test of the “emergence trauma hypothesis” developed from isolation studies on dogs (see above).
The isolates were “inferior” to controls in sexual, emotional, and dominance behavior. However, one wonders to what extent these differences resulted from factors other than total isolation. The isolates differed from one or both control groups in several ways other than being in total isolation for six months. These differences involved access to real mothers, access to peers, and separation from peers.
Sackett (1966) kept newborn rhesus infants in isolation chambers for nine months. The aim was not to induce abnormal behavior, but to see how “socially naive” infants would respond to pictures of monkeys. The pictures depicted monkeys of various ages and sexes engaged in different activities. The infants responded in a species-typical manner to certain pictures, and this was taken as evidence for unlearned or inborn responses to social cues. The abnormalities of these ‘picture isolates’ were described in later reports (Sackett 1967a, 1968a, b).
Follow-up Studies
Numerous follow-up studies were conducted on monkeys that had been reared in partial or total isolation at the University of Wisconsin. Several of these studies described the maternal behavior of females who had been deprived of their own mothers throughout infancy (e.g. Harlow & Seay 1966, Suomi 1977a). Harlow termed these females “motherless mothers.” These females had been reared in wire cages either alone, with cloth surrogates, or with peers. They were so disturbed by their deprived upbringing that few were impregnated without experimental intervention. Most of them were artificially inseminated or impregnated by males while restrained in a ‘rape rack’ — a device designed and named by Harlow.
All four motherless mothers in the first study were “totally inadequate” in caring for their first offspring (Seay, Alexander & Harlow 1964). Two infants died from neglect or abuse, one after becoming blind and partially paralyzed and the other after being refused mothers’ milk. All of the infants probably would have been killed if the experimenters had not intervened.
Most of the motherless mothers ignored their infants, but other motherless mothers abused their babies by crushing the infant’s face to the floor, chewing off the infant’s feet and fingers, and in one case by putting the infant’s head in her mouth and crushing it like an eggshell (Harlow et al. 1971b, p. 545).
(See Photo 8). Surprisingly, the motherless mothers in the Seay et al. study behaved more or less normally with their second infants.
 Photo 8
Photo 8
Typical behavior of an isolation-reared female towards her infant. The mother is crushing her baby against the cage floor. From H. F. Harlow, 1962, “The Heterosexual Affectational System in Monkeys,” Am. Psychol. 17:8. Copyright (1962) by the American Psychological Association. Reprinted by permission of the publisher.[/caption]
Despite neglect and abuse, infants of motherless mothers persisted in attempts to cling to, and suckle from, their mothers. In a rare display of sympathy, Harlow wrote:
One of the things that would unnerve experimenters was to watch the desperate efforts of these babies to make contact with the abnormal mother. She would beat them and knock them down; they would come back and make contact; the mothers would rub their faces into the floor; they would wriggle free and again attempt contact. The power . . . and demandingness of the infant to make contact and the punishment the infant would accept would make strong men reach the point that they could hardly bear to observe this unmaternal behavior (Harlow 1963, p. 25).
The social development of these infants was adversely affected by their traumatic upbringing (Arling & Harlow 1967). They “were deficient in social play and sexual behavior and hyperaggressive in peer interaction” (p. 371).
Several factors influenced the adequacy (or inadequacy) of maternal care by motherless mothers (Ruppenthal et al. 1976, Suomi 1978). These factors included (1) mother’s age at first social contact, (2) mother’s previous experience with peers or their own infants, and (3) sex of infant. For example,
… females with minimal . . . contact [with their previous infants] rarely showed maternal care improvement even after as many as five births. In most of these cases these females were hyper-abusive, either killing their baby soon after birth or mutilating it . . . (Ruppenthal et al. 1976, p. 345).
Gross ineptitude, not hyperaggression, was behind much of this abuse. For example, a female was seen “calmly chewing on its [infant’s] digits” and trying to “rip the remainder of the umbilical cord out of the infant’s midsection” (p. 346). However, more than ineptitude was involved. Abusive mothers “sometimes sought out the baby to strike or bite it without provocation” (Harlow et al. 1966, p. 59).
Other follow-up studies at Wisconsin examined different aspects of deprivation rearing. Sackett (1967a) found that the adverse effects of various deprivation experiences were persistent. He tested four-year-olds that had been reared in a variety of conditions, including solitary rearing in cages or isolation chambers and rearing with motherless mothers. The greater the deprivation experience, the greater the damage evident years later. The most deprived individuals were the most fearful and withdrawn and the least sociable and active.
Sackett also found that partial isolates and, especially, total isolates were less active and exploratory in novel environments than control monkeys (Sackett 1972a). However, female isolates were more normal than males on these measures, suggesting that in some respects, males are more vulnerable than females to isolation.
Sackett and coworkers conducted several studies of the social preferences of monkeys that had been reared in partial or total isolation (Sackett et al. 1967; Pratt & Sackett 1967; Suomi et al. 1970a; Sackett & Ruppenthal 1974). The aim was to relate these preferences to the isolates’ limited social experience or to their “innate” tendencies. Preferences were assessed in the “self-selection circus,” which had a central compartment that was separated by windows from several outer compartments. The chooser was placed in the center and the animals to be chosen were placed in the outer compartments. Preferences for particular “stimulus animals” were measured by the relative amounts of time the chooser spent near each candidate. Preferences were considered to be uninfluenced by the behavior of the stimulus animals, but this assumption was not necessarily true.
In one of these preference studies (Sackett et al. 1965], monkeys that had been reared alone in wire cages, but handled by humans during their first month, preferred humans to monkeys. The conclusion was “that the very early experience of a monkey with a specific type of stimulus can have lasting effects upon later preferential choice concerning that stimulus” (p. 306). Other results indicated that this very early experience can be overcome by later experience.
Sackett (1970a) concluded that the results of his preference studies do not support theories that emphasize the role of learning in the formation of social bonds. Sackett argued that monkeys are born with social tendencies, or they inevitably acquire these tendencies as they mature. These “response biases” lead to the acquisition of social attachments in the absence of learning opportunities. A second conclusion was that early rearing experiences are crucial in determining (1) “whether or not social attachments to species members will be formed” (p. 112) and (2) the nature of the resulting preferences (e.g. infants over adults). Although Sackett downplayed the importance of learning in the formation of social bonds, he believed that learning is important in the maintenance of social bonds.
The follow-up experiments that have been considered so far examined the maternal behavior and social preferences of maternally deprived monkeys. Follow-up experiments on these monkeys also investigated responses to stress. In one study, partial or total isolates did not differ from control monkeys in any hormonal measure associated with stress (Meyer & Bowman 1972). Monkeys were stressed by being placed in “primate restraint chairs” for two hours. Somewhat contradictory results were found in a second study (Sackett et al. 1973).
Habilitation Experiments
Wisconsin researchers made several attempts at habilitating monkeys reared in isolation (e.g. Chamove 1978). None of these attempts was motivated by sympathy for the disturbed monkeys. Rather, the rationale was to consider the “post-isolation syndrome” of rhesus monkeys to be a model of human mental disturbance, and to develop therapeutic techniques that may apply to humans.
The first attempted therapy simply involved giving isolation-reared monkeys opportunities to interact with one another. Partial isolates were moved to a zoo and placed on a monkey island that was surrounded by alligators (Harlow & Harlow 1962b, Harlow & Griffin 1965). This social experience had mixed results. “Fighting was severe at first” (Harlow & Harlow 1962b, p. 141). Two females were so injured that they were returned to the lab. One male drowned. Some friendships developed but sexual behavior remained impaired. The monkeys were returned to the lab, where friendly interactions ceased and fighting increased. “After two years we had to conclude that the island experience was of no lasting value” (Ibid., p. 141).
Sackett (1968b) attempted to habilitate socially withdrawn isolates by training them to approach one another. The isolates were placed in an apparatus with an electrified floor. The only way an isolate could escape shock was to jump onto a stool that already contained a monkey. The aim was to induce the monkey being shocked to join the monkey on the stool. This rather limited aim was met. However, the induced social contact failed to habilitate the monkeys.
The Wisconsin researchers eventually were successful in their efforts at habilitation (Harlow & Suomi 1971a; Suomi & Harlow 1972a; Suomi et al. 1972). Their subjects had spent their first six months in isolation chambers. Therapy involved pairing these isolates with ‘therapist’ monkeys (Photo 9). The therapists were younger and more normal than the isolates, but they too had been deprived of their mothers from birth. The isolates showed significant recovery in social and personal behavior. Success was attributed to the choice of therapists. These younger individuals sought contact without being threatening. They also could tutor isolates as they themselves developed social behaviors.
“Social therapy” was successfully extended to monkeys that had spent their first twelve months in isolation chambers (Novak & Harlow 1975, Novak 1979).
The success of the social therapy cast doubt on the Critical Period Hypothesis as an explanation for the devastating effects of total isolation on rhesus monkeys. This hypothesis states that certain social experiences facilitate behavioral development, but in order to do so, must occur during a critical period. If isolation prevents this, then the individual’s development should be permanently impaired. This does not appear to hold for rhesus monkeys.
Another possible explanation for the effects of isolation is the Emergence Trauma Hypothesis, derived from experiments on dogs (see above). Although emergence trauma may play a role in rhesus monkeys (Novak 1979), the Wisconsin researchers favor a learning deficit explanation (Suomi et al. 1974; Novak & Harlow 1975). According to this explanation, isolation precludes opportunities for social learning and leaves individuals severely disturbed. Under certain facilitating conditions, these disturbed individuals can learn species-typical behavior from more normal monkeys.
 Photo 9
Photo 9
Rhesus monkeys in a therapy experiment. On the left, a monkey reared in an isolation chamber. On the right, a ‘therapist’ monkey. Reproduced from Harlow (1974) with permission.[/caption]
Rhesus monkeys in a therapy experiment. On the left, a monkey reared in an isolation chamber. On the right, a ‘therapist’ monkey. Reproduced from Harlow (1974) with permission.
The process of social learning is extremely difficult because isolates are so withdrawn. Their self-directed behavior interferes with social interaction. Sackett (1970b) argued that isolates have difficulty inhibiting their self-directed behavior, as well as most existing behavior patterns. He proposed that this inability was responsible for the persistence of the isolates’ disturbance. Testing revealed that isolates do have difficulty inhibiting response patterns (Gluck & Sackett 1976, p. 174).
The success of the social therapy technique overshadowed the limited successes achieved by other techniques. These include exposure to cloth surrogate mothers (Suomi 1973a) and administration of drugs (McKinney et al. 1973; Nobel et al. 1976). In the latter study, one infant was so severely disturbed by the isolation procedure that “his disturbance behaviors included headbanging, convulsive jerks and spasms . . . and he seemed more susceptible to injury and illness” (p. 1169).
The long-term effects of both surrogate therapy and social therapy (involving ‘therapist’ monkeys) were assessed by Cummins and Suomi (1976). The habilitated monkeys showed no signs of regression, although the surrogate-habilitated group still exhibited only limited recovery.
Social Factors in Normal Development
Harlow and Harlow (1962b) discussed the development of rhesus monkey infants in various rearing environments. Their aim was to assess the ability of mothers and infants to facilitate normal development in infants. For example, Harlow and Harlow contrasted two social environments: mothers and infants versus cloth surrogate mothers and infants. Infant development in the two environments suggested that an infant’s relationship with its mother facilitates its social and sexual interactions with peers. Hansen (1966) reached a similar conclusion.
Various rearing experiments suggested that interactions with peers (other infants) were important for infant development. For example, Harlow and Harlow reared infants with one another, but not with mothers. They appeared to develop normally in their play and sexual behavior, despite the absence of real mothers. Harlow and Harlow suggested that interactions among infants “may compensate for lack of mothering” (p. 146). A similar conclusion was reached by other Wisconsin researchers (e.g. Hansen 1966, Sackett 1968a, Suomi & Harlow 1971).
However, later experiments provided less cause for optimism about the fate of peer-reared monkeys (Chamove et al. 1973). Peer-reared monkeys developed abnormally, perhaps because they clung excessively to one another. Moreover, their abnormality seemed to be persistent; for example, males often displayed incomplete mounting patterns and females were as abusive to their first-born infants as were isolation-reared females (Suomi & Harlow 1977a).
Miscellaneous Deprivation Experiments
Several deprivation experiments conducted at Wisconsin do not readily fit into the categories discussed above. These miscellaneous experiments are discussed in this section.
Sackett (1965a) presented a theory to account for the effects of early social deprivation on rhesus monkey behavior. According to this theory, normal behavioral development proceeds by gradual exposure to increasingly complex environmental stimulation. Animals are said to prefer stimulation that is a little more complex, but not much more, than what they are used to. Inappropriately paced exposure to environmental complexity is thought to result in some of the behavioral abnormalities of deprived monkeys.
Sackett conducted indirect tests of his theory (Sackett 1965b), using monkeys that had been subjected to varying degrees of isolation in previous studies. These tests offered admittedly limited support for the theory. The strongest conclusion consistent with the results is that animals reared under stimulating conditions prefer to interact with complex stimulation, whereas animals reared under impoverished conditions prefer simple stimulation. This conclusion received further support in follow-up testing (Sackett 1972a).
Kerr et al. (1969) conducted a statistical study of the physical growth of monkeys subjected to partial and total isolation in previous experiments. The isolates grew normally during and after isolation. The experimenters related their findings to a human syndrome known as “deprivation dwarfism” (see Discussion).
Lichstein and Sackett (1971) examined the reaction of isolation-reared monkeys to “noxious stimulation.” Isolation-reared monkeys and socially-reared monkeys were subjected to electric shock from the drinking spouts in their cages. The isolation-reared monkeys were found to tolerate higher levels of shock than the controls. The same monkeys were later given a choice between an electrified and non-electrified drinking tubes. Neither group preferred the electrified tube. This suggested that electric shock was a noxious stimulus for both groups.
Lichstein and Sackett concluded that rhesus monkeys reared in isolation react anomalously to noxious stimulation. The same phenomenon had already been shown in dogs and rats, but according to Lichstein and Sackett, the special feature of their study was that one explanation for the anomalous reaction, namely, that the stimulation was not perceived as noxious, was ruled out.
University of Pittsburgh School of Medicine
Researchers at the University of Pittsburgh conducted follow-up experiments on rhesus monkeys obtained from Wisconsin, where the subject had been reared in total isolation. The isolates were poor at sending social cues via facial expressions, and poor at utilizing cues sent by others (Miller et al. 1967). This was taken as support for Mason’s hypothesis that the social inadequacy of isolation-reared monkeys stems from lack of opportunities to acquire communication skills. This experiment subjected monkeys to electric shock and restraint in a primate chair for up to 72 hours.
Later experiments documented the eating and drinking habits of the isolates. The isolates consumed more food and fluids than socially-reared monkeys (Miller et al. 1969). This was attributed to a greater consumption of food and fluid during each meal, rather than, for example, more frequent meals (Miller et al. 1971). No reason was apparent for this phenomenon.
US. Public Health Service
Several studies of rhesus monkeys reared in partial isolation were conducted by the U.S. Public Health Service (PHS). Two results contrasted with the Wisconsin data. First, partial isolation had little or no effect on adult reproductive performance (Meier 1965a). Second, all ‘motherless mothers’ exhibited normal maternal behavior after normal delivery of their offspring (Meier 1965b). However, maternal behavior was inadequate following surgical delivery.
The contrast between the Wisconsin and PHS data was decreased when a later study (Missakian 1969) found that isolation did impair reproductive performance. Sexual behavior remained disoriented following social therapy, although some animals showed improvement in other aspects of social behavior (Missakian 1972). Therapy involved pairing isolates with younger individuals, as in the Wisconsin studies.
California’s Regional Primate Research Center at Davis
Mitchell and others at Davis conducted several deprivation studies on rhesus monkeys. Mitchell had conducted similar experiment as a student of Harlow’s.
Monkeys reared in isolation at Davis did not drink excessively, unlike those at Pittsburgh (Fittinghoff et al. 1971). One possible explanation, not mentioned by the researchers, is that polydipsia occurs in total isolates (Pittsburgh), but not partial isolates (Davis).
The Davis researchers made follow-up observations on a group of isolate monkeys obtained from Wisconsin (Fittinghoff et al. 1974). The 12 to 13 year-old partial isolates retained the abnormal personal and social behavior that characterized their younger years. The isolates’ abnormalities were viewed as normal responses to an aberrant environment “. . . the study provides few surprises by comparison with previous work . . .” (p. 136).
Baysinger et al. (1972) compared the behavioral development of mother-reared and isolation-reared infants during the rearing period. Both groups were housed in wire cages, but the isolates’ cages were covered. Unfortunately, the researchers removed the coverings during observations and made the dubious assumption that the isolates’ behavior was not affected.
The Davis researchers tried their hand at “social therapy” after the Wisconsin group successfully habilitated isolate-reared rhesus monkeys. Therapy involved pairing isolates with older males and females. “Although some of the isolate-reared animals appeared to have benefited from pairings with preadolescents [Brandt & Mitchell 1973], the isolates clearly gained little of lasting value” (Erwin et al. 1974, p. 814).
Capitanio (1984) recently conducted a deprivation experiment at Davis under the direction of Mason who — like Mitchell— had conducted deprivation experiments as a student of Harlow’s. Capitanio compared the social behavior of rhesus monkeys that had been reared by either a dog or a plastic hobbyhorse. The aim was to demonstrate that the animate, interactive characteristics of mothers enhances an infant’s social abilities. Dogs, but not hobbyhorses, have these characteristics. “Although members of both groups were clearly abnormal in the extent of their social interactions,” the dog-reared individuals were less aberrant than the hobbyhorse-reared individuals (p. 35).
Other deprivation experiments conducted at Davis examined the “looking behavior” and social organization of isolates (Mitchell 1972, Anderson & Mason 1974), and the affiliative behavior of an adult male isolate toward an infant (Gomber & Mitchell 1974).
Washington Regional Primate Research Center
Sackett, yet another of Harlow’s former students, continued his deprivation experiments at the Washington Primate Center after leaving Wisconsin. He began using pigtailed monkeys, a close relative of rhesus monkeys, expecting that pigtails and rhesus would react similarly to total isolation. Much to his surprise, Sackett found that “rhesus monkeys, as a species, appear to be more susceptible to isolation rearing effects than pigtails” (Sackett et al. 1976a, p. 128). Moreover, exposure to age-mates appeared to be an effective therapy for pigtails but not rhesus (Sackett et al. 1976b).
The results of a second study also emphasized species differences (Sackett et al. 1981). Rhesus and pigtailed monkeys, as well as the closely related crab-eating monkeys, were reared in total isolation. Effects on personal behavior, social behavior and exploration varied depending on species. The results cast doubt on the notion of an isolation syndrome and the generality of this syndrome.
Yerkes Regional Primate Center
Several deprivation experiments were conducted at the Yerkes Laboratories in Florida, and their successor, the Yerkes Regional Primate Center in Georgia. Only one Yerkes study involved rhesus monkeys. in that experiment, isolation, apparently total, “did not grossly affect basic learning abilities” (Mason & Fitz-Gerald 1962). Other studies all involved chimpanzees.
Davenport, Menzel, and Rogers conducted a series of deprivation experiments designed to address human clinical problems. For example, Davenport et al. (1 961) considered isolation-reared chimpanzees to be models of inadequate maternal care in humans. In humans, such care had been associated with poor growth and survival. The chimps were reared in bare wire cages or small isolation cubicles, and their growth and survival were compared to those of mother-reared chimpanzees. Rearing in wire cages was said to mimic rearing in impersonal human institutions. No explicit analogy was made for rearing in isolation chambers. Unfortunately, food intake—an important determinant of growth—was not controlled from group to group. The chimp data were contrary to the human data. Deprived chimps grew faster and survived better than nondeprived chimps. Harlow’s group reached the same conclusion in a later study (Kerr et al. 1969) and did not even mention the Yerkes study. These studies are discussed further in the Discussion section.
The Yerkes group also reared chimpanzees in different degrees of restriction. The most restricted infants were reared in bare “cribs” whose walls precluded visual access to the outside world. Other restricted groups were given more opportunities for visual, social, or manipulatory experience. Infants from these restricted groups were compared to infants that had been born in the wild.
The restricted infants were abnormal in several respects. First, they developed unusual stereotyped behavior and postures that persisted for years (Davenport & Menzel 1963). Second, they were unresponsive or fearful of objects (Menzel et al. 1963a) or unfamiliar environments (Menzel et al. 1963b). Third, they were slow learners, but improved with training (Davenport & Rogers 1968; Davenport et al. 1969; Rogers & Davenport 1971; Davenport et al. 1973). Their poor performance stemmed from inflexibility and “distractibility.” These results are consistent with human studies but inconsistent with rhesus monkey studies (see above). Fourth, sexual performance at adulthood was impaired initially, but recovered when isolates were aired with normal age-mates. Again, the chimp data do not agree with the rhesus data.
Some of this research was repeated using older chimpanzees. One to two-year-old infants were imported from Africa and subjected to six months of deprivation in isolation cribs (Davenport et al. 1966). Unlike chimps subjected to such procedures at birth, they did not develop stereotyped behavior. The researchers concluded that some as-yet-unknown forms of maternal stimulation, if provided early, can prevent stereotypy in chimpanzees later subjected to isolation.
Another deprivation experiment by the Yerkes researchers was a failed attempt to habilitate the isolation-reared chimpanzees (Turner et al. 1969). The subjects were adolescent chimps who avoided social contact and displayed abnormal behavior. These aberrations persisted despite various manipulations, including social therapy (pairing isolate males with same-aged females or older females), and drug therapy (including LSD).
Social therapy apparently was successful in another experiment (Rogers & Davenport 1969). Isolates were grouped with normal, same-aged males and females and showed marked improvement in sexual performance. The researchers claimed that recovery extended to social behavior in general, but unfortunately this was not documented.
The same chimpanzees reared in isolation by Davenport et al. were also studied by Berkson and Mason, who focused on the isolates’ abnormal stereotyped behavior. Stereotyped behavior was prominent in situations that induced excitement or “arousal”, as expected, and such behavior was reduced in situations that elicited alternative behavior (Berkson et al. 1963; Berkson & Mason 1964). One of these studies involved food deprivation and administration of amphetamines.
Another group of Yerkes chimpanzees was reared in partial isolation for two to four years and then tested in mildly stressful situations. Isolates gave fewer “distress vocalizations” (whimpers and screams) than wild-born chimps (Randolph & Mason 1969).
Delta Regional Primate Research Center, Louisiana
Some of the deprivation experiments conducted at Delta concerned self-recognition.The capacity for self-recognition was judged from the subjects’ responses to their reflection in a mirror. Unlike chimpanzees reared in the wild, those reared in partial isolation showed no signs of self-recognition, except when given remedial social experience (Gallup et al. 1971). The researchers argued that the self concept is developed through personal interactions, which are precluded by isolation. See Gallup and McClure (1971) for a related study on rhesus monkeys.
Another set of experiments compared the development of rhesus infants reared with surrogate mothers that were either mobile or stationary. Only the “stationary infants” developed stereo-typed body rocking (Mason & Berkson 1975). This was regarded as experimental support for human studies that suggested an association between maternal stimulation and body rocking. The “mobile infants” were less aroused by novel situations (Mason & Berkson 1975, Eastman & Mason 1975) and were more socially responsive (Anderson et al. 1977) than the “stationary infants.”
Miscellaneous Institutions
Researchers at numerous institutions conducted one or two deprivation experiments on nonhuman primates. For convenience, these experiments are grouped here, rather than under separate institutional headings.
Green examined some effects of partial isolation on rhesus monkeys. Compared to controls, the isolates were less exploratory and more fearful and hyper-reactive to environmental changes (Green & Gordon 1964). This interfered with their responses on learning tasks (Green 1965). Similar results were found in the Wisconsin and Yerkes studies.
Angermeier et al. (1967) reared rhesus monkeys in various degrees of deprivation and paired them with socially-reared monkeys in “dominance fights.” Isolates were not subordinate to the controls, unlike the situation in rats. The researchers concluded that studies of social behavior may yield different answers depending on the species being investigated.
Several experiments conducted at miscellaneous institutions involved species other than the popular rhesus monkey. Studies of partial isolation were extended to pigtailed monkeys (Evans 1967), cebus monkeys (Elias & Samonds 1973), and bonnet monkeys (Rosenblum & Smiley 1984). All three species resembled rhesus monkeys in their response to isolation. Some of the isolation-reared bonnet monkeys recovered substantially when they were required to gather their own food from a foraging device. The success of this therapy depended on the monkeys’ status in the dominance hierarchy. Kawabe (1969) reared Japanese monkeys in isolation, but he did not specify whether isolation was partial or total. In any event, his analysis was restricted to vocalizations.
Coelho and Bramblett (1984) reared yellow baboons in partial isolation except that each was allowed to interact with peers for a few hours per day. This arrangement was said to be subtly different from rearing with mothers and peers. This amazing assertion may have some validity, however, given that few differences in social behavior were found between the two groups.
Berkson continued his experiments on abnormal stereotyped behavior when he moved from Yerkes to the Illinois State Pediatric Institution (see Berkson 1967). Crab-eating macaques, but not marmosets, developed abnormal stereotyped acts when reared in partial isolation (Berkson et al. 1966; Berkson 1968). The macaques isolated earliest in infancy developed the most stereotypies.
Kaplan deprived squirrel monkey infants of their mothers and reared them with inanimate surrogate mothers. The infants formed an attachment bond to the surrogates and used them as a security base, much like surrogate- reared rhesus monkeys. However, unlike rhesus, the squirrel monkey infants did not exhibit stereotyped body rocking (Kaplan 1974).
Huebner and colleagues also reared squirrel monkeys with inanimate surrogate mothers. Surrogate-reared infants developed behavioral abnormalities, but recovery was substantial when infants were allowed to interact with a kitten (Huebner & King 1984). The kittens were thought to be important in providing a source of stimulation contingent upon the monkey’s behavior. “The importance of stimulation contingent upon an infant’s own behavior has been emphasized in human child development” (p. 241). The researcher noted that, for squirrel monkeys, this stimulation did not have to come from another member of the same species.
Several physiological experiments have been conducted on primates reared in isolation. In one experiment, Reite and Short (1977) examined the sleep physiology of pigtailed monkeys reared in total isolation. No differences from controls were found. Two other physiological experiments were based on the clinical observation that a biochemical abnormality in human schizophrenics was related to a monotonous rearing environment. Beckett considered the Wisconsin rhesus monkeys and the Yerkes chimpanzees to be animal models of humans reared in monotonous environments. Biochemical data from both rhesus (Beckett et al. 1963a) and chimpanzees (Beckett et al. 1963b) provided support for the link between the biochemical abnormality and a monotonous upbringing.
Another physiological experiment (Beasley & Seal 1968) was also related to schizophrenia. Partial isolation in rhesus monkeys was associated with alterations in the levels of several blood constituents. However, these alterations did not resemble those found in human schizophrenics. Indeed, many of the alterations were the opposite of those found in humans.
Discussion
The Nature and Extent of Suffering
Clearly, deprivation researchers have inflicted a great deal of damage on their animal subjects. Some animals were killed by the deprivation experience. Virtually all of the survivors suffered long-term psychological disturbance. For example, monkeys reared in isolation were social misfits, fearful and withdrawn, encapsulated in a world of abnormal stereotyped behavior and self-abuse.
The deprivation experience itself was not the only source of damage to the animals. A few of them were killed after their isolation. Many isolates were subjected to stressful procedures, such as electric shock and the “rape rack.” Because the isolates were so disturbed, virtually any procedure was distressing. Hence, the isolates were extremely fearful when exposed to novel environments, or were helpless when paired with aggressive individuals.
The damage inflicted by some experiments extended to the offspring of the participants. Because the maternal behavior of female isolates was grossly abnormal, many infants were neglected or abused; some died as a result.
These consequences of deprivation experiments were endured by over 1,300 animals. This figure includes 1,110 primates and 202 dogs. These figures are minimum estimates; many more animals actually were used. Unfortunately, numerous reports failed to mention the number of animals used in particular experiments.
Financial Cost
The estimated total cost of deprivation experiments was $20.7 million (Table 1). Most of this money was provided by the National Institutes of Health and the National Institute of Mental Health. Both organizations are federal institutions; hence, U.S. taxpayers ultimately paid for much of this research.

Benefits to Humans
Only one deprivation experiment has had a tangible impact on clinical practice. This was Suomi and Harlow’s (1972a) habilitation of rhesus monkey infants that had been reared in isolation chambers. Recall that these infants were habilitated during interactions with younger, more normal ‘therapists.’ This technique “has been applied successfully to treatment of socially …withdrawn children as well as those at high risk for schizophrenia” (Suomi 1982, p. 268).
The irony in this application of animal data is that the therapeutic technique could have been safely tested on humans, without using animals first. No potentially harmful drugs or surgical procedures were involved. Furthermore, the principles behind the success of the monkey habilitation were already familiar to psychiatrists. In discussing the habilitation, Suomi et al. (1972) observed.
The role of gentle . . . contact and model-serving exhibited by the therapists was likely of …importance for . . . recovery. Many of these principles are not unfamiliar to human psychotherapists (p. 931).
Clinicians had simply overlooked the potential therapeutic value of pairing certain kinds of disturbed children with younger, normal children. Suomi and Harlow’s contribution was to suggest this possibility.
Let us consider one of the two human studies that Suomi cited as having used his habilitation technique. This was a study of socially withdrawn preschoolers at a day care center (Furman et al. 1979). These children were labeled “social isolates” by Furman et al. and “socially retarded” by Suomi. Both labels are suggestive of a serious behavioral disorder; however, according to Furman et al., these children
were simply in the low end of peer interactions, compared to their classmates . . . none was described by the teachers as mentally retarded or emotionally disturbed (p. 917).
Hence, shyness in preschoolers was compared to the profound disturbance in rhesus monkeys following isolation-rearing.
These shy preschoolers became more sociable during interactions with younger individuals. The reason for this change was that the “isolates” became leaders in interactions with the younger individuals. The reverse was true in the monkey case. Even Furman et al. conceded that . . . the analogue between this study and the animal model . . . can easily be overstated . .” (p. 922). Accordingly, it can be argued that Suomi’s social therapy technique did not generalize to this human situation, and that the success of human study was serendipitous. Moreover, numerous techniques for overcoming shyness in human children already exist.
Suomi claimed that his habilitation study has had general theoretical impact in addition to alleged practical impact (Suomi & Harlow 1978, p. 268; Suomi 1984a). Specifically, the study was said to have had
considerable impact on theoretical considerations of developmental plasticity and the potential reversibility of early cognitive and social deficits at the human level . . . (Suomi 1984a, p. 232).
This assertion was unsupported. No mention of such an impact was made in numerous recent books on human development (see Reference Note, p. 40).
Aside from Suomi’s habilitation experiment, other clinically-oriented experiments reviewed have had little or no clinical impact. At best, some have added support to pre-existing human data or have suggested that more attention be paid to certain social factors, such as peer interactions.
Mental Illness in Humans
The limited impact of isolation experiments on the diagnosis, treatment, prevention, and understanding of human mental illness stems partly from the vagueness of the isolation model. The researchers usually failed to specify what human syndrome they were modeling. They talked vaguely of “mental disturbance” (Harlow & Harlow 1962a), “mental illness” (Mitchell 1974), “abnormal behavior” (Sackett 1968a), and “psychosis” (McKinney 1974a). Each of these terms refers to a heterogenous assortment of human clinical syndromes. Accordingly, no single animal model for any of these assortments is likely to go far. With respect to mental illness, the post-isolation syndrome remains an animal model in search of a human counterpart.
Even Mason, himself a Wisconsin researcher, believed that the use of animal models of human behavior works best for well-circumscribed processes, not for example, “the sources of neurotic behavior” (Mason 1968, p. 71). Sometimes the isolation researchers did draw parallels to specific human problems, such as autism (e.g. Fox 1966), depression (e.g. Griffin & Harlow 1966), and schizophrenia (Harlow et al. 1964; Sackett 1972b). Some analogies were more convincing than others, but no researcher has yet demonstrated that the post-isolation syndrome is a good animal model (both valid and practical) of any human mental illness. Such a demonstration is unlikely given that the response to isolation varies both within and across animal species (e.g. Fuller & Clark 1968; Meier 1970; Sackett et al. 1976).
Maternal Deprivation in Humans
Recall that isolation experiments were viewed as animal models of maternal deprivation as well as mental illness. Both the Wisconsin and Yerkes researchers explicitly related their isolation experiments to the clinical literature on maternal deprivation.
The term ‘maternal deprivation’ was used rather loosely in the human literature (see Ainsworth 1962). It referred primarily to an insufficient level of maternal care. Such insufficient care prevailed in non-nurturing orphanages, foster homes, and families. Numerous clinicians reported that children reared in impersonal institutions and other depriving environments are prone to develop abnormally (Bowlby 1952). Some of these children, but by no means all, develop emotional, intellectual, social, or physical problems.
Some of the early human studies were criticized for methodological or conceptual inadequacies (e.g. Yarrow 1961, Andry 1962). Some studies were criticized for ignoring non-maternal variables. For example, deprivation of sensory stimulation as distinct from maternal care —appeared to be a contributing factor in the deprivation syndrome (Casler 1961). This was especially so for infants less than six months old, which is the period before an infant becomes emotionally bonded to its mother. However, the significance of maternal deprivation, especially for infants over six months of age, was apparent from the human studies (Ainsworth 1962, Lebovici 1962, Thompson 1962, Bronfenbrenner 1968).
The isolation experiments that came closest to mimicking conditions in impersonal institutions for children were (1) the Wisconsin experiments with rhesus monkey infants housed alone in wire cages (‘partial isolation’) and (2) the Yerkes experiments with chimpanzee infants similarly housed. The Wisconsin cage-reared monkeys, insofar as they resembled children being reared in impersonalized institutions, provided support for the negative effects of maternal deprivation in humans (Bowlby 1976, Mussen et al. 1984). The same can be said for the Yerkes chimpanzees.
Harlow attempted to relate his total isolation experiments to human maternal deprivation (Harlow & Harlow 1962a; Harlow et al. 1965), but his attempt was far from convincing. A panel of the National Academy of Sciences concluded that total isolation comes closest to modeling cases of drastic neglect in human infants that probably would lead to death (Hamburg 1970). Monkeys reared in isolation chambers are deprived of all social experience, maternal or otherwise. Moreover, total isolates are deprived of normal sensory experiences. Hence, isolation effects could stem from lack of sensory experience, as well as maternal deprivation per se. Yarrow (1961, p. 480) noted that even “in the most extreme institutional environments the degree of sensory deprivation is less severe than in animal studies.”
It is ironic that Harlow did not control for sensory experience in his total isolation experiments. Diminished sensory experience had been a criticism of some of the early human studies (Caster 1961). Harlow was fond of criticizing clinical studies for their lack of experimental rigor, and he touted his own ostensibly well-controlled experiments. Harlow et al. (1964) dismissed sensory experience as a confounding factor by claiming that “. . the monkeys [inside the chambers] had constant stimulation, . . . including contact stimulation from manipulation of their [cage’s] iron bars . .” (p. 120). This meager level of stimulation clearly did not approach the level that monkeys usually experience in the wild or even in the laboratory.
These were not the only problems with the total isolation model of maternal deprivation. Sackett’s group found that the effects of total isolation vary across monkey species:
The results . . . suggest a word of caution concerning generalization of behavioral effects of isolation rearing across species . . The generality of rearing condition effects, even across very closely related non-human primate species [cannot be] simply assumed (Sackett et al. 1976a, p. 129).
These results cast considerable doubt on Harlow’s belief
…that the same basic laws operate for [human and rhesus infants] and that social isolations which produce abnormality in one species will have comparable effects on the other (Harlow et al. 1965, p.90).
A decade after Harlow’s initial experiments on total isolation, he reassessed the contribution of these studies to the understanding of human maternal deprivation (Harlow & Novak 1973). His extremely weak assessment was couched in terms of his trumpeted experimental control.
The information we have obtained . . . differ[s] from human data only in the fact that the precise controls which we could exercise over our monkey subjects make the differential concepts [of privation and deprivation] plausible and precise (p. 467).
Again:
. . . our data give clarity and classification to the earlier clinical efforts (p. 477).
A decade before Harlow’s isolation experiments, Ainsworth and Bowlby (1954, cited by Lebovici 1962) distinguished between deprivation of the mother from birth (privation’) and deprivation after an infant developed an emotional bond to its mother (‘deprivation’). The differential effects of privation and deprivation were so striking in the human studies that no animal studies were necessary to add plausibility or precision to the human data. Moreover, Harlow’s experiments actually confounded privation and deprivation. The monkeys subjected to privation were all incarcerated in isolation chambers at birth, whereas the monkeys purportedly subjected to deprivation were not incarcerated until six months of age. However, even the latter monkeys were subjected to privation because they spent their first six months without their mothers. The inescapable conclusion is that Harlow’s total isolation experiments added nothing to our under-standing of maternal deprivation in humans.
It is puzzling that Wisconsin researchers came to view their isolation experiments (partial or total) as models of human maternal deprivation. The earliest isolation studies at Wisconsin were said to be investigations of what was already known for humans: “deprivation of normal social experiences in human children results in a wide range of personal and social deficiencies and aberrations . .” (Mason 1960, p. 582). The rationale for Mason’s experiments was that no comparable data existed for non-human primates. Sackett (1967b) also acknowledged prior human studies. After summarizing the effects of various deprivation experiences on rhesus monkeys, he wrote:
a wealth of clinical data and data on institutionalized children suggests that many of the conditions discussed in this paper have similar, if not identical, effects in human children (p. 75).
One of few isolation experiments to go beyond existing human data suggested that isolation-rearing does not impair physical growth or survival, at least in chimpanzees (Davenport et al. 1961). Later studies on humans (Whitten et al. I 968) and rhesus monkeys (Kerr et al. 1969) yielded similar results. These studies suggested that “deprivation dwarfism” in institutionalized children results from malnutrition, not social isolation. However, the conclusions of both animal studies were tempered by methodological flaws.
The biggest breakthrough in maternal deprivation research apparently was the realization that impersonal institutions and other depriving environments can impair child development. Bowlby (1952) called attention to this problem back in 1952, when he reviewed the existing human data. His monograph was largely responsible for a marked improvement in institutional care for children (World Health Organization 1962, p. 7-8) The Wisconsin and Yerkes data came after this.
To summarize: Deprivation experiments made little contribution as animal models of maternal deprivation. Many of these experiments were little more than sensational illustrations of what had already been established in humans.
Development in Human Infants
The Wisconsin researchers sought to justify their deprivation experiments not only as models of mental illness and maternal deprivation, but also as models of infant development. Harlow and Harlow (1962a), for example, aimed to help “resolve the baffling complex roles of various kinds of early experience in the development of human personality” (p. 146).
The Wisconsin researchers apparently were not successful. In 1976, Suomi wrote:
. . . whether actual data obtained from nonhuman primates have added measurably to our understand-ing of human development is another matter The record to date has not been particularly impressive. To be sure, there are a few findings derived from nonhuman primates that have clearly advanced knowledge of human development [Cites affection experiments]. Overall, however, such cases are relatively rare. Most monkey data that readily generalize to humans have not uncovered new facts about human behavior; rather, they have only verified principles that have already been formulated from previous human data.
A case in point is in the area of the development of mother-infant relationships. To date the monkey data have added little to knowledge of human mother-infant interactions (Suomi 1976, p. 203).
A year later, Suomi (1977b) implied that Wisconsin research has influenced research in child development.
The importance of monkey peer relationships for social development has been stressed for years (e.g. Harlow 1969); only after many years of disinterest have human development researches [sic] directed more than passing attention toward the development of infant and childhood friendships . . . (p. 222).
The importance of peer relationships among monkeys was emphasized in a viewpoint that Suomi (1979) offered to students of human infant development. Suomi provided several examples of how this viewpoint — derived from his rhesus monkey experiments — might be used to plan social policy:
[This] viewpoint would emphasize greater focus on the study of peer interaction patterns in children and it would encourage a policy of providing children with ample opportunities to develop mutual relationships in stable and non-threatening environments. It would argue that peer relationships in younger children are unlikely to thrive if problems with parents (and perhaps teachers and other adults) remain substantial, leading to prevention programs that focus on individuals’ entire social environment, not simply their relationships with peers or with parents. Finally, such a viewpoint would argue for careful monitoring of children’s relationships with peers as a barometer of their current level of social competence and as a possible predictor of subsequent psychopathology (p. 146).
These policies may well be beneficial. However, to base such policies on Wisconsin’s monkeys seems unwise, if not absurd. All of these monkeys are admittedly abnormal (Harlow, in Cullen 1974, p. 422; Suomi 1977b, p. 212) and living in highly artificial conditions.
There is a more specific reason to be skeptical. One of the most important features of any model of human development is the nature of mother-infant interactions. Here the rhesus monkey model falls short. According to the Wisconsin researchers, play is one of an infant’s most important activities; yet these researchers acknowledge that “rhesus mothers engage in little interactive play with their infants, . . . unlike human mothers” (Harlow & Harlow 1969, p. 21, emphasis added). Given this model, perhaps it is not surprising that the Wisconsin researchers have emphasized the role of infant-infant interactions over that of mother-infant interactions. Consider Suomi and Harlow’s (1975a) conclusion:
. . . the primary vehicle for shaping genetically acquired potential into competent and adaptive social activity lies in the development and maintenance of peer friendships [among infants] . . . we suspect it is also . . . true for . . . man” (p. 176).
While this suspicion may be true, differences between the model and humans do not inspire confidence.
The Wisconsin researchers have tried to get students of normal social development in humans to consider the implications of their rhesus monkey data (Mason 1968, Suomi 1979). For example, Suomi and Harlow (1978) offered two implications: (1) a child’s environment can have significant effects on behavioral development and (2) these environmental influences are particularly important at the beginning of various maturational stages. Suomi and Harlow proclaimed these implications as if they were novel, but of course they were not. Indeed, the first (the importance of a child’s environment) is regarded as a truism. Beach and Jaynes (1954) reviewed numerous experiments documenting the importance of early experience in animals. All of these were conducted well before Harlow’s research. Back in 1952, Levy wrote that the importance of early experience in humans was recognized by “psychiatrists generally, child psychologists, educators, and a variety of investigators in other disciplines” (p. 489). Levy added that the importance of early experience is “easily verified by the study of case records in any number of schools and clinics” (p. 489). In his view, experiments to “document” this are attempts “to verify the obvious” (p. 489). Hence, the Wisconsin researchers weren’t the first to document the importance of early experience; their contribution was to develop a new model for research on this topic (Hamburg 1970).
Suomi and Harlow’s second implication is that environmental influences become particularly important at the beginning of certain maturational stages. This had already been shown, for example, by early human studies of maternal deprivation (reviewed in Ainsworth 1962). One such study (Ainsworth & Bowlby 1954, cited in Lebovici 1962) distinguished between deprivations occurring before and after the infant forms an emotional attachment to the mother. This occurs at around 6 months of age, and at this maturational stage the consequences of deprivation seem most severe.
While primate deprivation models have yielded numerous implications about human development, these implications are largely ignored by clinicians, who rely primarily on human studies. A few implications from deprivation experiments have filtered into clinical thinking, but these do not appear to have influenced clinical practice. Only one recent book out of several that were consulted even mentioned an assessment of the clinical impact of the Wisconsin models of normal infant development (Reference Note, p. 40). Referring to experiments by Harlow and others, Kolb and Brodie (1982) wrote:
From animal studies it has been inferred that the establishment of the . . . relationship [of an infant to its mother] comes about through the presence of the mother, whether her behavior may be judged as gratifying or thwarting, rewarding or punishing (p. 69).
This statement provides little evidence that Harlow’s models of infant development made significant contributions to the understanding of human development.
Child Abuse in Humans
Recall that in the Wisconsin experiments, female rhesus monkeys who were reared without mothers often became inadequate mothers themselves. The poor maternal behavior of motherless mothers has been mentioned in the psychiatric literature by Kolb and Brodie (1982, p. 69) and White (1974). White went so far as to say that the “demonstration of the intergenerational effects of deprivation [has] had a significant influence on psychiatric thinking . .” (p. 18). It is difficult to imagine the basis for this glowing assessment. Would White have been surprised if human females who had been reared in isolation became poor mothers?
Suomi has related the Wisconsin deprivation experiments to human child abuse (e.g. Suomi 1977a). His assessment has been cautious. For example, he evaluated the primate model according to McKinney and Bunney’s criteria for a valid animal model (see Ch. 2) and judged the model to be weak:
The existing data on monkey neglect and abuse of offspring scarcely begin to satisfy all of these criteria (Suomi 1978, p. 32).
Hence, there is little basis for expecting that preventative or therapeutic measures that work for rhesus monkeys will work for humans.
Suomi recently summarized Wisconsin data on infant abuse and drew several conclusions (Suomi & Ripp 1983). First, several factors contribute to the problem of infant abuse. These include mothers’ social history and infants’ demandingness. Second, mothers who were the most adversely affected by particular social experiences are likely to be the most abusive. Third, therapy/prevention can be helpful by reducing exposure to risk factors and giving special attention to high-risk females. None of these conclusions is likely to come as a surprise to students of human child abuse.
Horenstein (1977) was more sanguine than Suomi regarding the potential of Wisconsin experiments to contribute to clinical aspects of child abuse. Horenstein faulted fellow clinicians for ignoring the Wisconsin data. He and others distinguished “motherliness” (physical contact, warmth, emotional involvement, etc.) and practical mothering (feeding, cleaning, etc.). He claimed that monkey infants who were denied motherliness, but given practical mothering, developed deficits in their own motherliness. This is incorrect; these monkeys were denied practical mothering for all but the first few weeks of their lives. Throwing monkey chow into a cage hardly counts as practical mothering (breastfeeding, grooming, etc.). Moreover, a link between deprivation of motherliness as a child and deficits in motherliness as a mother has been shown by human studies, as Horenstein notes.
Horenstein thought that early Wisconsin experiments suggested two treatment programs: supervised mothering for abusive mothers, and peer interactions for abused children. He conceded that both types of treatment were developed independently of Harlow’s work, but he pointed out that some clinicians do not treat both infants and mothers in their programs.
Horenstein also claims that theoreticians, unlike clinicians, regarded Harlow’s work as important to understanding child abuse. He cited Steele and Pollack (1968) and Fontana (1973) to support this claim. However, Horenstein exaggerated these theoreticians’ assessments of Harlow’s work. Consider Steele and Pollack’s assessment. After quoting Harlow, these authors write:
These observations do suggest that there may be some connection between the lack of early “being mothered” experience and deficits in later parental functions (p. 117).
That is the only statement regarding the positive contribution of Harlow’s work. The authors had previously noted that human studies (not to mention common sense) revealed the same connection between early experience and later maternal behavior. The authors then go on to point out differences between Harlow’s monkeys and abusive parents. They conclude by suggesting that Harlow’s monkeys are more like infants reared in depriving institutions, rather than like mothers prone to abuse. Hence, Steel and Pollack’s paper provides little support for the claim that Harlow’s work made important contributions to the understanding of child abuse.
The same conclusion holds for Fontana’s book. Fontana referred to the monkeys reared on cloth surrogate mothers and those reared on wire surrogate mothers (see Ch. 3 above). The former were said to develop more normally than the latter, who, denied any semblance of maternal affection, sometimes developed into abusive mothers. Fontana then added “Similarly, the indifference of a human parent can be shattering to a child . .” (p. 57, emphasis added). In other words, what we already know about abusive human mothers also appears to be the case in rhesus monkeys.
Duplication
What is the extent of unnecessary duplication of experiments in deprivation studies? A reasonable assessment of unnecessary duplication is possible by examining the goals, results, and number of studies in this field.
Consider the premises upon which the following assessment is based. First, most of the deprivation experiments that we have considered were directed at human problems. These experiments were not designed, or funded, to amass knowledge for its own sake. For example, Harlow designed his research to address human problems (Harlow, in Tavris 1973). Most of the money for deprivation research was supplied by the National Institutes of Health, whose “mission . . . is to protect and improve the health of the American people” (National Institutes of Health 1984, p. iii).
A second premise is that deprivation experiments that were not justified on the basis of human benefits should not have been conducted, given their damaging effects on animals. A third point concerns the argument that no animal experiment, however poorly conceived, is unnecessary because it can serendipitously yield important results. Research on animal subjects should follow the same ethical principle that applies to research on human subjects; namely, an experiment is either justified or unjustified at the outset, regardless of outcome (Levine 1985). Self-serving appeals to the vagaries of luck are not excuses for bad research.
Deprivation studies had several goals; most of these studies were attempts to document the effects of isolation, the persistence of these effects, and their experimental reversal. In many cases, all three of these aspects of isolation could have been documented using the same subjects.
Over 175 reports of deprivation experiments have been published through 1984. To be sure, each of these reports does not represent a separate study; 36 contain reviews of experiments published in other reports. This leaves 146 studies, nearly all of which involved primates (Table II).

The obvious question is why so many studies were conducted. The 146 studies do not comprise the same type of study conducted on species after species; most studies involved rhesus monkeys. Rather, they comprise study after study of different aspects of the deprivation experience. Various kinds, durations and consequences of deprivation were examined. For example, Harlow and colleagues subjected monkeys to total isolation 14 times and devoted part or all of 21 follow-up studies to total isolates. The differences among several of the original 14 studies involved the timing and duration of isolation. The relevance of any of these studies to humans is suspect, given that human infants probably would perish if subjected to total isolation (Ham-burg 1970). The total isolation studies appear to provide an example of what Rowan (1984) labels “parametric tinkering.”
Research Quality
Several aspects of deprivation research are open to question on scientific grounds. As we have seen, the experimental design of many of these studies was seriously flawed. Harlow himself criticized numerous deprivation experiments for faulty design. His target was research on the association between deprivation and learning ability. He all but dismissed the studies that did not involve primates. For example, he wrote:
The subprimate literature possesses a wealth of massive . . . and often meaningless researches .. . The primary contribution of these earlier researches is the illustration of errors in experimental design and procedure (Harlow et al. 1971a, p. 126).
Harlow et al. thought these errors led researchers to the wrong conclusion (p. 147).
A second scientific aspect of deprivation experiments that is open to criticism is the choice of research questions. At the time of the primate experiments, the adverse consequences of rearing human infants in relative isolation, without mother love, were well established. The more controversial issue of the day was what aspects of maternal care facilitated infant development at various stages. Specifically, was the general sensory stimulation provided by mothers more important than emotional aspects of mother love? Rather than concentrate on this question (or similar issues), the animal modelers played it safe by “experimentally demonstrating” the ill effects of maternal deprivation.
A third criticism of deprivation research is that dogs and other nonprimates are not good choices for animal models. The dissimilarity to humans is too great. No evidence has been found that any of the experiments on nonprimates has influenced clinical thinking or practice. Although nonhuman primates were better models, the excessive reliance on one species (rhesus monkeys) was poor research strategy. The relatively few studies of monkeys other than rhesus monkeys demonstrated substantial variability in response to deprivation from species to species. This variability cast doubt on the generality of the results from these studies.
Alternatives
The issue of alternatives is almost moot with respect to deprivation experiments. It does not apply to the bulk of deprivation experiments because of the following ironies. First, animal data on the effects of maternal deprivation came after the human data (see above). Second, insofar as the animal data related to mental illness or child abuse, it was of dubious value (see section on clinical impact). Even some of the modelers themselves conceded that the models violated criteria of validity. Third, some of the animal data, though possibly interesting, made little difference to human affairs.
Only one deprivation experiment had a direct clinical impact, and as already mentioned, the procedure derived from that experiment could easily have been derived from human studies.
One of the themes of the Wisconsin deprivation experiments is the importance of peer interaction for infant development. Here again, animals did not have to be deprived of their mothers to determine this. The important comparison was not monkey infants reared only with peers versus only with mothers. (No one would suggest that human infants be reared by their peers instead of their mothers, regardless of the outcome of the monkey experiment.) The relevant comparison was mother-peer rearing versus mother-only rearing. In this perspective, alternative studies with humans are readily apparent and undoubtedly have already been conducted. Researchers can compare the development of mother-reared infants who have various degrees of contact with peers. Infants could be given enhanced levels of peer contact, or researchers could take advantage of naturally occurring variability in peer contact, such as differential enrollment in day care programs.
Animal studies not only were unnecessary, in light of the accomplishments and potential of human studies, they also were excessively harsh. This translated into more damage to animals and less relevance to humans. Mason recognized this when he observed that deprivation studies can be “likened to the blow of a sledge hammer, when a few gentle taps might have been more instructive” (Mason 1968, p. 79). For example, total isolation studies were more damaging and, as has been argued above, less relevant to humans than were partial isolation studies. Similarly, animals did not have to spend their entire lives in isolation in order to demonstrate the importance of social experience in early life.
Summary
Experiments in the deprivation category involved rearing infant animals in various degrees of social isolation. The aim of the experiments was to determine the effects of this isolation on development.
Over 1,300 animals, mostly primates, were subjected to these experiments. Most suffered long-term psychological damage, and some were physically harmed or even killed. These experiments cost over $20 million. Most of this funding was supplied by federal institutions.
Although nearly 150 deprivation experiments were conducted, only one had a direct impact on clinical practice. The procedure involved in this study could have been easily derived from human studies, without prior testing in animals.
This field of research contains many experiments that were either poorly designed or unnecessarily duplicative of prior research, including human studies. Moreover, alternatives to many of these animal experiments were either already in use or could have been devised.
Reference Note
These books included Handbook for the Practice of Pediatric Psychology (Tuma 1982), Child Development and Personality (Mussen et al. 1984), The Child: Development from Birth Through Adolescence (Harris 1984), Social and Personality Development: Infancy Through Adolescence (Damon 1983), Child Development (Abron 1984), Modern Clinical Psychiatry (Kolb & Brodie 1982), and Handbook of Research in Early Childhood Education (Spodek 1982).
style=”text-align: right;”> style=”color: #808080;”>
Please cite as:
Stephens, Martin L. (1986) Maternal Deprivation Experiments in Psychology: A Critique of Animal Models.
Retrieved from http://aavs.org/maternal-deprivation-experiments-psychology.
Experiments in Separation
Like experiments in the deprivation category, separation experiments involve removing infants from their mothers. However, the timing of this removal differs between the two types of experiments. In deprivation experiments, the removal occurs before the infant becomes psychologically attached to its mother. In separation experiments, the removal occurs when the infant is older, after is has become attached to its mother. One other difference between separations and deprivations is that the former usually are temporary, whereas the latter are permanent.
There are several types of studies that are related to mother-infant separation. These involve separating infants from other infants, confining infants in vertical chambers (also known as “pits” and “wells of despair”), or rearing infants with punitive artificial mothers. These studies are far outnumbered by experiments involving mother-infant separation (Table Ill). All of these miscellaneous studies have been conducted on rhesus monkeys at the University of Wisconsin. These studies are considered here because of their historical and conceptual association with mother-infant separation studies; maternal separation experiments led to the other types of experiments and all of these studies ‘are considered to be animal models of depression.

Researchers have offered several reasons for conducting separation experiments on animals. Mother-infant separation has been considered to be an animal model of mother-infant separation of humans, as well as a model of any depression or anxiety that results from the separation.
‘Infant-infant’ separation studies have similar rationales. Studies involving vertical chambers or punitive surrogate mothers were attempts to create animal models of psychopathology.
Not all separation experiments on animals have been conducted as animal models of human phenomena. Specifically, a few of the early animal studies of mother-infant separation are best characterized as basic research.
Human Studies of Mother-Infant Separation
Primate research on mother-infant separation is based on human studies that began in the 1940s. Many of these pioneering human studies were the same ones that formed the basis of animal studies of maternal deprivation (see Ch. 4). Psychiatrists such as Bowlby, Spitz, and Goldfarb had been studying children in orphanages, hospitals, and other settings. They found that children sometimes protested being separated from their mothers and later lapsed into despair. Bowlby termed this the “protest-despair reaction” to separation. Spitz (1946) coined the term “anaclitic depression” for this form of despair. Some children also showed a third phase, detachment, in which they were indifferent, if not hostile, to their mothers during brief visits or when reunited.
Early human studies not only identified the protest-despair-detachment reaction to separation, but also began identifying factors that influence the reaction’s severity. The influence of factors such as the child’s age at separation, the availability of alternate care givers during the separation, and the duration of separation, were reviewed in two World Health Organization monographs (Bowlby 1952, WHO 1962). The early human research also began addressing the longer-term consequences of maternal separation on personality development.
Animal models of maternal separation came into use in the 1960s, two decades after the early human studies. The animal modelers advanced their usual criticism of human studies, namely, that these studies were not rigidly controlled experiments. The models purportedly were going to provide definitive answers using monkeys. The models were used to assess (1) the short- and long-term effects of a temporary separation from the mother, and (2) the factors that influence the severity of these effects. Both behavioral and physiological effects of separation were studied.
Summary of Animal Separation Experiments
Separation experiments have been conducted on nonhuman primates, dogs, cats, and rodents, with the primate studies accounting for about three-quarters of the total number of studies (Table III).
Cat and Dog Experiments
Three separation studies have been conducted on cats. These involved prematurely separating kittens from their mothers and sometimes their littermates as well (Seitz 1958, 1959; Rosenblatt et al. 1963; Rosenblatt 1974; Guyot et al. 1980). These studies apparently were designed as basic research, and are rarely (if ever) mentioned in the literature on separation models of human phenomena. Seitz’s study is noteworthy for its harmful effects. All the cats were stressed, some became “catatonic,” and some died.
Puppies were the subjects of several clinically oriented separation studies. Most of these studies were conducted by J.P. Scott and coworkers at the Jackson Laboratories. The separations in Scott’s experiments usually lasted only a matter of minutes, but this was enough to induce acute distress. Longer separations had far more damaging consequences.
Two of Scott’s most damaging experiments involved repeated, overnight separations or repeated 22-hour separations. The former was described as a “drastic emotional experience” accompanied by “serious losses in weight” (Scott & Bronson 1964, p. 183). In the latter, “the animals were . . . severely upset. . . . [They] ate little . . . and one . . . even died. . . . ” (Scott et al. 1973, p. 13).
The most noteworthy findings from Scott’s experiments were: (1) Separation from social companions induces fear or loneliness. (2) Separations are most damaging when they occur during the ‘critical period for socialization.’ (3) Various psychotropic drugs are not “effective in reducing distress vocalizations that are elicited by separation, except at doses that produce sedation or reach near-toxic levels” (Scott et al. 1973, p. 17). Scott and Bronson thought that their “most important result [was] the discovery of a general social drive,” (p. 192) based on fear or loneliness. According to the researchers, the existence of such a drive should be investigated in humans.
Scott and coworkers were not the only researchers to conduct separation studies on dogs. Senay’s (1966) study had an unusual twist: laboratory-reared puppies were separated from the experimenter, not their mother. (Mothers had been removed earlier). This study was an unsuccessful attempt to create an animal model of depression.
Rodent Experiments
Myron Hofer, a psychiatrist at a medical college, has been prematurely separating rat pups from their mothers since the late 1960s. His experiments have emphasized physiological as well as behavioral variables. He gave two rationales for his experiments (Hofer 1970). First, little attention had been given to effects of mother-infant separation in “lower” mammals such as rats, and second, few studies examined the physiological effects of separation.
Hofer attempted to systematically analyze the pups’ separation experience into its component parts. Specifically, he viewed the separation experience as consisting of the loss of a source of nutrition, warmth, tactile stimulation, olfactory stimulation, etc., as well as the psychological loss of the mother. Hofer attempted to relate these aspects of the separation experience to the physiological and behavioral responses to separation. Once a particular cause and effect relationship was indicated, Hofer attempted to determine the biological mechanism underlying this relationship.
In a typical experiment, Hofer would “cull” his rat litters to 8-10 pups each, implant electrodes in the pups, remove the pups from their mothers, and monitor certain physiological and behavioral changes during the next 24 hours. The fate of the pups after this period is unclear from Hofer’s reports.
The separated pups usually were divided into groups that were subjected to different treatments. Some treatments functioned to control for certain variables. For example, one separated group might receive milk through a stomach tube as a control for maternal nutrition. Another group might be housed with an unrelated adult female who potentially could provide all forms of stimulation that a mother could (e.g. tactile stimulation), except providing milk. The same goal was sometimes achieved by returning the mother to the separated pups after her teats were ligated.
Hofer’s experiments had two themes. First, different aspects of the maternal separation can have different effects, each mediated by a different biological mechanism (Hofer 1973a). Second, rat mothers are regulators of physiological processes in their pups (Hofer 1978).
In Hofer’s (1970) first experiment, maternal separation caused a drop in heart rate and respiratory rate. These responses were related to deprivation of nursing (Hofer & Weiner 1971; Hofer 1971, 1973b). Mother’s milk regulated infants’ heart rate via a neural pathway, not the circulatory pathway (Hofer 1973a). The experiment that showed this involved cutting the pups’ spinal cord in various locations. Follow-up experiments (Hofer & Weiner 1975) involved a host of surgical and biochemical procedures.
Separated pups were hyperactive when placed in a novel environment (Hofer 1973c). This was related to the loss of mothers’ warmth, as well as mothers’ tactile and olfactory stimulation (Hofer 1973a, b, c, d; 1975a, 1976a). These experiments involved (1) simulating tactile stimulation with electric shock and (2) destroying the pups’ sense of smell with chemicals. Not surprisingly, the hyperactive pups were also slow in falling asleep (Hofer 1973b). Deprivation of maternal nutrition was implicated in this response.
The pups’ sleep-wake cycle was radically altered by maternal separation (Hofer 1976b). This experiment was highly invasive, involving implantation of cranial electrodes and head plates. Hofer noted that other researchers had obtained similar results in rats, as well as in guinea pigs and monkeys.
Fourteen day-old pups did not survive permanent maternal separation unless supplied with heat (Hofer 1975b). Even with heat, over 50% of the pups died. All separated pups initially showed decreased physiological functioning. The survivors showed a physiological recovery that was attributed to physical maturation.
Hofer and coworkers (Stone, Bonnett & Hofer 1976) performed one of the most damaging series of separation experiments. In survival experiments, pups were permanently deprived of their mothers and some were also denied food and water for a week. Time of death was noted. In metabolic experiments, pups were killed and their bodies “were minced and dried . . . in [an] oven” (p. 243). In developmental experiments, pups were decapitated before or after a starvation period.
The topic in this study was the role of body temperature in survival and development. One separated group was supplied with warmth and another was not. No pups lived through the survival experiments when denied food, though the warm pups survived a little longer than the pups without warmth. When food was available, no pups denied warmth survived whereas most pups with warmth survived. After summarizing their results, the experimenters noted that “similar consequences of prolonged hypothermia have been observed in . . . human infants, young rabbits, and piglets . . . ” (p. 246).
In another experiment, Hofer and coworkers subjected pups to stressful procedures in order to induce stomach “erosions,” It was already known that maternal separation increases pups’ susceptibility to these erosions. The follow-up experiment (Ackerman, Hofer & Weiner 1975) found that this susceptibility was age-dependent. The procedure involved subjecting starved rats to restraint. Some pups developed “massive hemorrhage.” The mechanism for the susceptibility to gastric erosions was later found to be faulty temperature-regulation, induced by maternal separation (Ackerman, Hofer & Weiner 1978).
Researchers other than Hofer conducted separation experiments on rodents. Ader and co-workers conducted three separation studies that were part of a series of experiments linking early social experiences to later susceptibility to stressful or pathogenic procedures. The first of these three experiments was the basis for Hofer’s experiments on gastric erosions (see above).
Ader’s experiment linked early maternal separation to susceptibility to gastric erosions (Ader et al. 1960) All pups were killed either by the experimenters or by the procedures themselves. Ader’s second experiment suggested that the increased susceptibility to gastric erosions does not result from loss of mother’s milk (Ader 1962). In his third experiment, Ader subjected pups to injections of cancerous cells (Ader & Friedman 1965). The separated pups showed higher mortality than non-separated ones.
Newell (1967) subjected mice to early maternal separation and then tested their ’emotional reactivity’ and learning ability. Based on human studies, he expected ’emotionality’ to be high and learning ability to be low. No such effects were found. Not only did the mice respond differently than humans, the two strains of mice reacted differently from each other.
Plaut and Davis (1972), two psychiatrists working at a hospital, conducted a series of mother-litter separations in rats. A high percentage of the separated pups died. A lower percentage died if the pups were kept with a non-lactating adult female (“aunt”). The same improvement in survival was also achieved by returning mothers to their pups after the mothers’ nipples had been burned shut. The only important difference between these findings and Hofer’s earlier studies (see above) was that Plaut and Davis showed that the aunts interacted with the pups.
Another experiment by Plaut and Davis (1972) concerned the effect of premature weaning on body weight and brain chemistry. This experiment also overlapped heavily with previous experiments cited by the authors.
Plaut later repeated some of his experiments using “uncles” as well as “aunts” (Plaut et al. 1984). Uncles, like aunts, reduced the high death toll of early weaning. The mere presence of an aunt behind a screen also had a positive effect on pup survival, though not as great as when aunts could interact with pups. Post-mortem examinations linked death to high urine retention, although experimental stimulation of urination failed to reduce pup mortality. Koch and Arnold (1976) performed a follow-up experiment to Hofer’s demonstration that heart rates in rats drop following maternal separation (see above). The researchers knew from Hofer’s work that artifical feeding would ameliorate this change in heart rate and that the presence of a non-lactating foster mother would not. The researchers wanted to know if foster mothers would have an ameliorating effect while pups were artificially fed. It did. To show this, the researchers surgically excised the teats of foster mothers to preclude nursing.
Primate Experiments
Separating primate infants from their mothers can have profound consequences. As we shall see, it can induce anxiety, despair, and even death. The act of separation and as well as the period of maternal absence can be stressful. Separation is often accomplished by brute force. One study noted that separation required “a trained team of three men” (Seay & Harlow 1965). The following quote describes the immediate response of pigtailed monkeys to the separation procedure:
Separation of mother and infant monkeys is an extremely stressful event for both mother and infant, as well as . . . for all other monkeys [nearby]. The mother becomes ferocious toward attendants and extremely protective of her infant. The infant’s screams can be heard almost over the entire building. The mother struggles and attacks the separators. The baby clings tightly to the mother and to any object to which it can grasp to avoid being held or removed by the attendant. With the baby gone, the mother paces the cage almost constantly, charges the cage occasionally, bites at it, and makes continual attempts to escape. . . . The infant emits . . . shrill screams intermittently and almost continuously for the period of separation (Jensen & Tolman 1962a, p. 132-133).
Another quote in the same report describes the reunion of mother and infant:
The reuniting of mother and infant is a remarkable event. Both return to each other immediately. The mother sits quietly holding her baby, and if no attendants are present, she very quickly seems content and relaxed. All is quiet in the room. No more piercing screeches of the baby or sounds from the mother are heard. . . . [After 15 to 40 minutes,] the infant would venture away from its mother and appear to be free to leave and explore as it did in the control situation, or, it would go to sleep being held by the mother . . . (p. 133).
University of Wisconsin and Wisconsin Regional Primate Center
Separation experiments at Wisconsin initially involved mother-infant separation, but later also involved infant-infant separation, vertical chamber incarceration, and miscellaneous procedures. All of these experiments were conducted on rhesus monkeys, as were the Wisconsin experiments on affection and deprivation.
Seay, Hansen, and Harlow (1962) began the Wisconsin separation experiments to see if the human “protest-despair-detachment” reaction (see above) also occurred in rhesus monkeys. For unexplained reasons, Seay et aL separated mothers and infants by putting a transparent partition between them. Therefore, mothers and infants could still see and hear each other, unlike the human situation. One infant in this study “suffered severely from anorexia and sleeplessness” (Harlow and Harlow 1971, p. 218). In a follow-up experiment (Seay & Harlow 1965), mothers and infants were separated visually as well as physically.
Similar results were obtained in both experiments. Not surprisingly, the infants initially were highly agitated, crying and struggling to reach their mothers. When their agitation subsided, they cried and played little with other infants in an adjoining cage. Mothers and infants eagerly embraced one another after their three-week separation was terminated. Within a few weeks their relationship seemed to return to normal.
Seay et al. interpreted their subjects’ reaction to separation as one of protest and despair; they therefore concluded that their results were in “general accord” with human data. They admittedly found little evidence in rhesus monkeys for the detachment phase.
Mitchell, Harlow, and coworkers (Mitchell et al. 1967) separated infants from foster mothers as well their own mothers. Infants were briefly but repeatedly separated from one mother until transferred to another. These manipulations left the infants psychologically disturbed. The disturbance was evident a year later when these youngsters, as well as others in control groups, were paired with an aggressive, fear-provoking adult male.
Suomi et al. (1970b) introduced infant-infant separations as an alternative procedure to mother-infant separations. They noted that separating infants from other infants had certain methodological advantages over separating them from their mothers. The act of separating infants from each other was easier to accomplish. Because of this, infants could be easily separated from one another repeatedly, and more data on transient responses to separation could thereby be obtained. These advantages brought one disadvantage, however, in that infant-infant separation has less relevance to mother-infant separation in humans.Suomi et al. therefore proposed infant-infant separation as a general model for studying loss of a loved one or ‘attachment object.’
Suomi et al. separated a group of infants from one another no less than twenty times. During their four-day separations, the infants were kept in physical — but not visual — isolation. No explanation for this research design was provided. The reaction to separation was similar to the protest-despair response to mother-infant separation; however, the repeated separations were so traumatic that they stunted the infant’s behavioral maturation.
Separation studies at Wisconsin and elsewhere were viewed in a new light after McKinney and Bunney (1969) called for development of animal models of depression. More attention was paid to the depression-like despair phase of the response to separation. Thus, mother-infant separation and infant-infant separation were considered animal models of depression.
Harlow created the infamous vertical chamber as another means of modeling depression (Harlow & Suomi 1971b). Harlow himself described this chamber as “a modified form of sadism” (Harlow, in Tavris 1973, p. 76). This tiny chamber had sides of stainless steel that sloped inward (Photo 10). This design frustrated the attempts of incarcerated monkeys to climb up the sides and cling to the wire mesh roof. After a few days of scrambling around, incarcerated monkeys spend most of their time huddled on the floor of the chamber (Suomi & Harlow 1972b, see Photo 11).
 Photo 10
Photo 10Vertical chambers, also known as ‘pits’ and ‘wells of despair,’ used to induce psychological disturbance in monkey infants. Reproduced from Harlow (1974) with permission.
The devastating effects of vertical chamber confinement were revealed in two studies (Harlow & Suomi 1971b, Suomi & Harlow 1972b). Thirty and 45 days of incarceration both produced “severe psychopathology.” After confinement, the infants spent most of their time huddling and clasping themselves, and performing other self-directed “disturbance behaviors.” “Most striking was the virtual absence of social activity” during months of social testing (Suomi & Harlow 1972b, p. 14). Suomi and Harlow saw “enormous potential” for the vertical chamber, in that its severe effects were produced rapidly and lasted indefinitely. Harlow had managed to create a device that was more devastating than isolation chambers (McKinney et al. 1971).
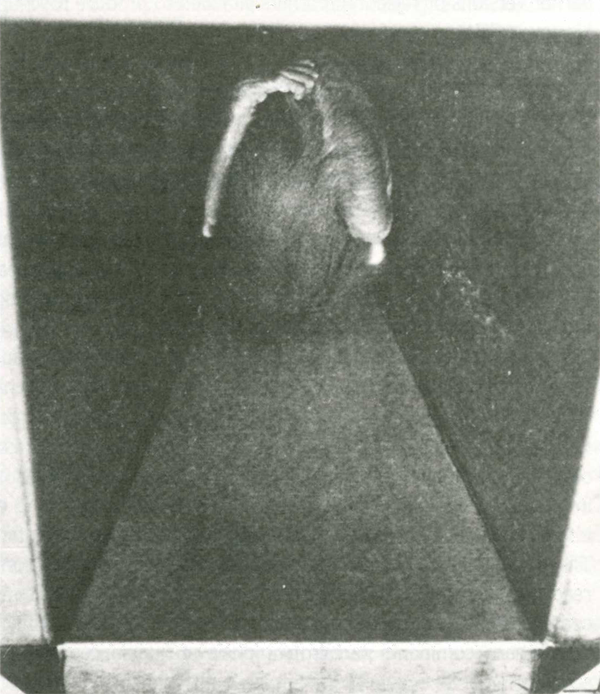 Photo 11
Photo 11Typical behavior of a rhesus monkey infant during long-term confinement in vertical chambers. The view is from the top of a chamber. From Harlow (1976) with permission.
Harlow and Harlow (1971) summarized some vertical chamber experiments that remain unpublished in detailed form.
McKinney, who had called for development of animal models of depression (see above), later joined forces with Harlow and Suomi at Wisconsin. McKinney and his new coworkers presented their studies of infant-infant separation and vertical chamber confinement as attempts to “go beyond the mother-infant separation model” of depression (McKinney et al. 1971, p. 134). McKinney’s praise of vertical chamber studies is puzzling. These studies radically violated one of his proposed criteria for an animal model to be valid; namely, that a syndrome be produced in the animal model by the same or similar cause of the syndrome in humans (McKinney & Bunney 1969). No one would argue that depression in humans is caused by vertical chamber confinement.
The Wisconsin researchers devised another odd method of inducing depression. Infants were reared with cloth surrogate mothers whose bodies could be made ice cold, too cold for the “punished” infants to cling to (Harlow et al. 1973). The cold surrogate was considered an advance over previous attempts to build a rejecting surrogate mother. These earlier versions were either made of uncomfortable material (e.g. sandpaper), or could dislodge infants with hidden spikes, catapults, compressed air or vigorous shaking (Baysinger et al. 1973). These earlier versions of rejecting mothers all failed to produce psychopathology in infants (Harlow & Harlow, 1971).
In the Harlow et al. study, brief drops in the surrogate’s temperature failed to induce depression. Harlow then kept the surrogate cold throughout the night, when infants invariably cling to their surrogates. The infants’ responses were so severe that one died. This was termed “psychological death.” All infants were severely dehydrated. “Although we feared for their lives, we did not discontinue the nocturnal cold . . . period until . . . we received the necropsy report on the dead infant” (p. 305).
According to Harlow’s article, he continued to subject infant monkeys to this procedure; however, no further publications resulted from these experiments. He considered this cold-surrogate procedure to be a model of extreme depression, though no mention is made of this model in reviews of animal models of depression.
The separation and vertical chamber models of depression had a longer lifespan at Wisconsin than did the cold-surrogate model. In fact, the separation and chamber procedures were some-times combined in the same experiment (Suomi 1973b, Suomi & Harlow 1975b).
Lewis et al. (1976) abandoned mother-infant separation as a model of depression after obtaining disappointing results. The results were from a series of five experiments that varied in the degree of access that infants had to their mother (and others) after separation. Most infants were killed after several days of separation, and no infant was returned to its mother. The results were highly variable and did not support the notion of a clear-cut protest-despair reaction. Lewis et al. thought that “future research in separation models of depression should concentrate on the peer separation paradigm” (p. 705). It should be emphasized that the researchers’ comments pertain to models of human depression, not models of mother-infant separation per se.
Two Wisconsin experiments (McKinney et al. 1973b, Young et al. 1973) used “well-controlled experiments” to test a well-known finding of human clinical psychology. The clinical finding was that the effects of stressful situations in adulthood are more severe if such situations were experienced during childhood. McKinney et al. (1973b) employed vertical chamber incarceration as their stressful procedure. Monkeys were profoundly disturbed by vertical chamber incarceration, but their disturbance was no greater if they had been subjected to similar treatment earlier in life.
The early stressful experience in the second study was separation from peers, coupled with vertical chamber confinement. The later stressful experience was no less than 28 separations from peers, one per day and 23 hours each. The experimental group did have a more severe reaction to the multiple separations than did a control group that was not subjected to early peer separation and chamber confinement. “Our data indirectly support previous studies in human subjects … ” but these data “are best viewed as tentative . . . and suggestive of further research” (p. 404). The mixed results of these two studies shed little light on the question of early behavioral sensitization in rhesus monkeys, let alone humans.
Most Wisconsin separation studies examined the effects of temporary separations from mothers or peers. Suomi et al. (1973) examined the effects of permanent separations from mothers. The severity of the short- and long-term responses to separation depended on (1) the infants’ age at separation and (2) whether the infant was housed alone or with a peer following separation. “One is tempted to conclude . . . that the reaction to separation is intensified by subsequent individual housing but mollified by subsequent social housing, a conclusion reached long ago by Spitz (1946) from study of human infants” (p. 382, emphasis added). The researchers admittedly could not even draw this modest conclusion, however. They pointed out that the disturbance of the solitary-housed monkeys could have resulted from the restrictive housing and not the trauma of separation.
The number of novel separation experiments that can be designed is limited only by the imagination of the researchers. This point is well illustrated by a Suomi et al. (1976) study in which four infants were first separated from their mother by wire mesh, then by a plastic panel, and then were separated from each other by a transparent panel. A period of reunion with mothers and peers followed, and this, in turn, was followed by permanent separation from the mother. Infants in a control group were subjected only to the final, permanent separation. Reaction to the wire separation was relatively mild, but the two other temporary separations were “debilitating.” However, the behavior of these individuals eventually resembled that of the non-separated controls during the reunion period. Both groups reacted similarly, and relatively mildly, to the final, permanent separation, despite their different early histories.
The results were contrary to the researchers’ expectations. Suomi et al. expected increasingly severe reactions to the first three separations, and major differences between experimentals and controls in reaction to the final separation. The results are at odds with Young et al. (1973), who reported that early separations tend to exaggerate reactions to later separations. Numerous uncontrolled factors and defects in experimental design were offered to account for the discrepancies between studies.
In their 1977 review, Suomi and Harlow conceded that much of their depression research had not gotten beyond what was already known for humans. A decade and a half of their experiments had been devoted to validating the rhesus monkey model of depression. Later studies were also attempts to further validate the model. For example, Suomi et al. (1978) wanted to see if a particular anti-depressant drug already in use for humans also worked in depressed monkeys. The drug imiprimine lessened the severity of the response to peer separations, but its effectiveness waned rapidly after treatment was discontinued.
Lewis (1978) conducted a series of experiments that apparently have not been published except in abstract form. Three groups of infants were separated from their mothers and temporarily housed alone in wire cages or isolation chambers, or housed together in a peer group. Individuals in the group had the most severe reactions, which included “failure to thrive” (p. 4509). Lewis, following Cairns, attributed this response to the complexity of the peer group’s separation-environment, in which the loss of mothers as social facilitators was most acutely felt.
A 1983 report by Suomi et al. noted that no previous primate study had repeatedly separated mothers and infants over short periods of time. Their study accomplished this. They were interested in the short- and the long-term effects of repeated maternal separations on infants. Mothers and infants were separated eight times, reunited for several weeks, and then the whole process was repeated. A second group of infants was not separated from their mothers. Eventually both groups were permanently separated from their mothers and housed in peer groups. Infants protested after each separation but this waned with repeated separation. Signs of the despair reaction were minimal. However, repeated separations appeared to somewhat retard development of the mother-infant relationship. Suomi et al. discussed the similarities and differences between these findings and those of earlier Wisconsin studies. The patterns in protest and despair reactions were consistent with social learning theory.
Wisconsin researchers focused on the behavioral response to separation until the 1970s; thereafter, physiological as well as behavioral responses were examined. Most of these physiological/behavioral experiments were conducted by McKinney and coworkers in the Department of Psychiatry. These experiments sometimes involved invasive surgical procedures, even death, in addition to the usual psychologically traumatic procedures of earlier studies.
Physiological and behavioral data gave conflicting evidence as to whether infants formed stronger attachments to their mothers or to inanimate surrogates (Meyer et al. 1975). Strength of attachment was inferred from the infants’ response to stress during maternal separation. Stress was induced by placing infants with “mechanical monsters” that made loud noises and sudden movements.
Another physiological study centered on the protest phase of the protest-despair response to separation. Breese et al. (1973) wanted to know if the anxiety characteristic of the “protest response” was associated with the production of certain chemicals in the brain (catecholamines). Human and animal studies had already suggested a link between anxiety and catecholamine production. Breese et al. separated infants from their mothers, observed them in the protest stage, and then killed them to analyze their brains. As expected, catecholamine-producing chemicals were elevated.
A related study focused on the despair phase of the protest-despair response to separation (Kraemer & McKinney 1979). AMPT, a drug that inhibits catecholamine production, was administered to infants that were repeatedly separated from peers or mothers, or infants that were reared in isolation. The resulting dose-response relationships revealed that AMPT induced depressive effects in all groups, but that the peer-separated group was more sensitive to this effect than was the mother-separated group.
This experiment was based on clinical reports linking catecholamine inhibitors to depression in a small percentage of human patients. Clinical data suggested that these patients shared certain predisposing factors. The rationale for the animal study was as follows:
While some of these factors, such as previous history of depression, ongoing psychosocial and environmental stress, can be intuitively identified, there are practical as well as ethical problems involved in actually testing the relative contribution of these factors in precipitating human depression and thereby determining their importance in a quantitative fashion (p. 33).
According to the researchers, the contribution of their animal experiment was in demonstrating that catecholamine inhibitors do interact with developmental and social stressors, such as peer separations.
In a recent study by McKinney’s group, cerebrospinal fluid (CSF) was extracted from monkeys undergoing repeated peer separations (Kraemer et al. 1983). The results indicated that monkeys with relatively low levels of norepinephrine in their fluid are: (1) “more likely to show a severe despair response to . . . separation” and (2) “more sensitive to the [catecholamine] depleting effects of AMPT and show greater depressant effects of the drug. Repeated separations do not affect the level of CSF [norepinephrine], but reductions in [other CSF components do occur]” (p. 141). These results suggest that low CSF norepinephrine is a stable characteristic of certain individuals, and predisposes them to severe responses to separation. In contrast, levels of other CSF components fluctuate in response to repeated separations.
The researchers viewed depression as an outcome of “life events,” such as separation, and constitutional factors, such as CSF norepinephrine levels. The existence of constitutional factors is one reason for individual differences in response to separations.
Recently, individual differences in response to separation also were emphasized by Suomi and coworkers. However, Suomi’s group focused on anxiety, whereas McKinney’s group focused on depression. Suomi et al. (1981) gave four reasons in arguing that anxiety was well-modeled by social separations: (1) the protest phase in the protest-despair response to separation resembles an anxiety-driven panic attack in humans; (2) rhesus and human infants display similar ‘anxious attachments’ to their mothers after reunion; (3) both display similar anxiety in anticipation of predictable separations; and (4) both display similar recovery after administration of the drug imipramine (Suomi et al. 1981).
The Wisconsin monkeys were considered to be anxious when they displayed behavior normally associated with fear, but in the absence of fear-provoking circumstances. This behavior was highly variable depending on the monkey’s age, sex, background, and current circumstances, but it included self-directed disturbance behavior (see Ch. 4) and excessive clinging to the mother. Monkeys with a history of separations displayed a chronically high level of anxious behavior. They appeared “edgy” or “uptight.” This high baseline level of anxiety was increased during “life threatening” events, such as an ongoing separation. However, a monkey’s background and current circumstances did not always permit an accurate assessment of its anxiety level. Constitutional factors, inborn or genetic, also appeared to be involved.
Following the human literature, Suomi’s group began looking for links between anxious behavior and the autonomic nervous system, which governs physiological variables such as heart rate and breathing rate. The responsiveness of the autonomic system to stressful situations was compared to levels of anxious behavior. The first measure of ‘autonomous reactivity’ to be investigated was heart rate (HR).
Month-old monkeys were repeatedly exposed to a mild stressor (a sound stimulus) and their change in heart rate (ΔHR) was monitored. Throughout one year of surrogate rearing, those infants with the highest ΔHR had the highest levels of anxious behavior. In contrast, ΔHR was not correlated with other behavior (Baysinger et al. 1978).
The same monkeys were monitored until puberty (Suomi et al. 1981). After their surrogate mothers were removed they were housed in partial isolation and subjected to various stressful procedures (e.g. daily handling). ΔHR values taken early in life continued to predict levels of anxious behavior. Hence, the researchers succeeded in linking individual variation in anxious behavior to a constitutional factor (ΔHR). Moreover, there was “suggestive” evidence that this factor was heritable.
Suomi (1981) conducted a “replication study,” this time using monkeys reared with peers rather than surrogates. The infants were subjected to repeated separations from peers during their second, fourth and tenth months. The findings were similar to those in the earlier studies. ΔHR was correlated with anxiety levels, but only during stressful conditions (during and shortly after separations).
The most recent study in this series (Suomi 1983) involved HR and a second measure of autonomic reactivity: plasma cortisol (a hormone associated with stress). Both measures were correlated with one another when the monkeys were exposed to stressful conditions (e.g. peer separations), but not under baseline conditions. Both physiological variables were correlated with anxious behavior measured months before or after the physiological recordings; again, however, the correlations were apparent only under stressful conditions.
The results suggested that individuals who have extreme behavioral reactions to stress also have extreme physiological reactions. These extreme behavioral and physiological reactions are stable over time, but highly reactive individuals appear “normal” in the absence of stress.
Suomi has suggested that excessive anxiety can be associated with two other problems, excessive depression and retarded social development. The same individuals who are highly anxious during the protest phase of separations are also the ones who become depressed if the separation endures (Suomi 1986). This claim awaits published supporting evidence, however. Suomi also argues that chronic anxiety inhibits exploration and positive social interaction, and thereby retards social development (see also Suomi & Harlow 1976).
University of Cambridge, England
Several mother-infant separation experiments were conducted at the University of Cambridge. The subjects were rhesus monkeys, as in the Wisconsin studies. However, the monkeys at Cambridge lived more natural lives than those at Wisconsin. They were housed as groups in large outdoor enclosures. The Cambridge researchers, under the direction of ethologist Robert Hinde, were therefore able to study aspects of mother-infant interactions, such as spatial relationships, that were largely precluded by the restrictive housing in the Wisconsin experiments.
In a typical Cambridge experiment, mothers were taken from their six-month old infants and returned six to 13 days later.
Infants in the Cambridge studies clearly were disturbed by separations (Hinde & Spencer-Booth 1966, Spencer-Booth & Hinde 1967). The separated infants usually sat in a hunched posture, crept about on their hind legs, and otherwise acted depressed. They gave “distress calls,” apparently to signal their mothers. The separated infants also were more susceptible than non-separated infants to mildly frightening tests, and paid less attention to new objects in their environment. The separated infants sought care from other individuals in the group, but received far less than their mother would have provided.
These infants were emotionally unstable when reunited with their mothers. They clung to their mothers and remained near them to an exaggerated degree. They threw “violent tantrums” when their mothers started to reject them. They were characterized as “anxiously attached” to their mothers.
The seriousness of the infants’ disturbance was evident in other ways. The infants sometimes became very upset for no apparent reason, or they awoke “terrified” and clung to their mothers. Also, signs of the disturbance persisted at least two years (Hinde & Spencer-Booth 1968; Spencer-Booth & Hinde 1971c, Hinde et al. 1978). Some of the testing for lingering disturbance involved subjecting infants to “mildly frustrating” situations, such as having food placed just outside of their reach or being threatened by a researcher wearing a “grotesque” mask.
A Cambridge study (Spencer-Booth & Hinde 1971a) had alarming results. Nineteen infants of various ages were separated. Of these, seven died during the experiment. For the youngest ones, the researchers were “unable to say how far the deprivation experience contributed to the deaths . . . ” (p. 179, emphasis added). Two other infants collapsed and were prematurely reunited with their mothers. The survivors responded to separation as in the first study, with depression during separation and excessive clinging and trailing of the mother during reunion.
The researchers concluded that the responses of rhesus monkeys and humans to maternal separation were similar. Their conclusion is convincing only for the despair stage of the three-stage human response (protest, despair, detachment). The evidence for protest in the Cambridge experiment was distress calling accompanied by much idle sitting. This idleness contrasts with the frantic scampering that other researchers liken to the protest reaction in humans. Moreover, there was no evidence of detachment in these studies.
These early Cambridge experiments all involved six-day separations. Spencer-Booth and Hinde (1971b) wanted to determine if rhesus infants would be more disturbed by longer separations. The researchers separated infants for thirteen days and compared the results to those from their earlier studies using one or two six-day separations. After reunion (but not before), “the severity and durability of the depression [were] related to the length of the separation” (p. 604). One infant died during this experiment.
One finding from the early Cambridge experiments was that rhesus monkey infants differed markedly from one another in the severity of their response to separation. The infants’ age, sex, and associations with other individuals accounted for little of this variation. A follow-up experiment related this variability to tension in the mother-infant relationship (Hinde & Spencer-Booth 1970). Infants who had the tensest relationships with mothers during reunion were the most disturbed. Tension was measured by the degree to which infants were rejected by their mothers and were responsible for maintaining proximity to their mothers. Tension after reunion was correlated with tension before separation. This indicated to Hinde and Spencer-Booth that rhesus mothers make only limited adjustments to individual differences in the demands of their infants after separation. The researchers conceded that in other species, such as humans, mothers might be more flexible in meeting the post-separation demands of their infants.
This experiment suggested that rhesus infants are disturbed not only by the separation itself, but also by the resulting disruption in their relationship with their mother. The same conclusion was reached by a later study (Hinde & Davies 1972a, b). This study, unlike previous Cambridge studies, involved removing infants, not mothers, to a strange environment during the separation period. It was expected that infants would be more disturbed in the “infant-removed” situation than in the “mother-removed” situation. The reverse occurred. This was attributed to the mothers’ behavior. Mothers were less disturbed when left in their home environment during separation, and therefore were better able to tend to their infants at reunion. In the mother-removed experiments, mothers at reunion had to divide their time between reestablishing dominance relations with other group members, and tending their infants. The researchers thought that there were no human parallels to these findings (Hinde & Davies 1972a, b).
A follow-up study (McGinnis 1978) supported the conclusions of the previous study. Both infants and mothers were removed from their social group and then separated them from each other. The results from this “mother and infant removed” study indicated that removal of mothers disturbs mothers, which in turn, causes them to be less responsive to their infants upon reunion. Mothers that ranked lowest in their group’s dominance hierarchy were most disturbed by removal from their group. Consequently, their relationship with their infants took longest to recover.
In another study, McGinnis (1979) removed both mothers and infants from their groups but not from each other. The results indicated that maternal separation, and not removal from group companions, was responsible for infant disturbance.
Downstate Medical Center, New York
Kaufman and Rosenblum conducted several mother-infant separation studies at Downstate Medical Center. Rosenblum is a former student of Harlow. His subjects were two species of monkeys closely related to rhesus monkeys: pigtailed monkeys and bonnet monkeys. The adult pigtails, and probably the bonnets as well, had been born in the wild. The monkeys were housed in groups in relatively large indoor cages. During the separation experiments, mothers were removed from these groups for up to several weeks. Their infants were several months old at the time of separation.
 Photo 12
Photo 12An agitated pigtailed monkey after its mother was taken away as part of a separation experiment. The infant is grimacing in fear while fleeing from an adult female that had just rebuffed its approach. Reproduced from Kaufman & Rosenblum (1969) with permission.
The first Downstate study (Kaufman & Rosenblum 1967a, b) showed that pigtail infants were severely disturbed by removal of their mothers. The act of separation elicited loud screams and involved a “massive struggle.” Infants were “acutely distressed” for the rest of the day on which their mothers were removed (Photo 12). The next day, three of the four infants were deeply depressed (Photo 13).
Each infant sat hunched over, almost rolled into a ball. . . . Movement was rare. . . . The movement that did occur [usually] appeared to be in slow motion. . . . The infant rarely responded to a social invitation or made a social gesture, and play virtually ceased. . . . Occasionally he would look up and coo (1967a, p. 1020).
The researchers interpreted these responses as fitting the protest-despair framework, only they referred to these phases as “agitation” and “depression”, respectively. A third phase, not seen in the Rhesus monkey studies, was “recovery.” Recovery was gradual and not complete even at the end of a month of separation. One infant showed only the agitation and recovery phases. It had turned to other adults for comfort during the separation period, whereas the other infants tried to cope alone. Only the latter became depressed.
 Photo 13
Photo 13A pigtailed monkey after its mother was taken away as part of a separation experiment. The infant is “showing the characteristic posture and the face of ‘grief.’ ” Reproduced from Kaufman & Rosenblum (1969, p. 687) with permission.
Mothers and infants showed a resurgence of interaction when reunited (Photo 14). Some aspects of the mother-infant relationship remained altered for at least three months, as in the Cambridge studies. These lingering effects contrast “dramatically” with the results of Seay and Harlow.
The most significant aspect of this study is that pigtails responded to separation in a qualitatively different manner from rhesus monkeys. As mentioned above, rhesus did not show the pigtails’ recovery stage. Moreover, neither species showed the detachment stage found in humans.
A more dramatic example of species differences was provided by the next Downstate study (Rosenblum & Kaufman 1968, Kaufman & Rosenblum 1969). The researchers repeated their earlier study, but used bonnet macaques instead of pigtails. The striking result was that the depression phase was virtually absent. The lack of depression was attributed to the care that separated infants received from other adults. Indeed, some infants were fully adopted, although all but one returned to its mother at reunion. These studies showed that data from one animal species do not necessarily generalize to a closely related species—let alone to humans.
The Downstate researchers subsequently made various attempts to induce depression in bonnet infants. One scheme was to remove mothers and leave infants with unresponsive foster mothers. Another scheme was to remove infants from their mothers and place them alone in a cage. Both procedures failed to induce noticeable depression. Kaufman attributed these negative results to the infant’s age and maturity. These results were briefly summarized by Kaufman (1973) but have yet to be published in detail.
 Photo 14
Photo 14A pigtailed monkey mother and infant reunited after a separation experiment. Reproduced from Kaufman & Rosenblum (1969) with permission.
Kaufman eventually succeeded in making bonnet infants depressed. His experiment (Kaufman & Stynes 1978) was conducted after he moved from Downstate to the University of Colorado Medical Center. A bonnet infant was reared in a group that contained pigtails as well as bonnets. When the bonnet infant reached five months of age, the researchers removed its mother and all other bonnets from the group. The infant exhibited pigtail-like severe depression.
The same report also detailed two other sets of experiments; these duplicated Kaufman’s earlier studies of pigtails and bonnets. The only noteworthy difference in the follow-up experiments was that bonnets showed more indications of depression than in the earlier studies, though the reaction still was not severe.
The most recent report of separation experiments at Downstate was in 1980 (Rosenblum & Plimpton ‘1980). Pigtailed infants looked depressed during the first separation night, but not during the daylight observations the next few days. Rosenblum and Plimpton viewed the infants’ behavior both as a response to loss of mother and as an attempt to cope without the mother. According to this view, infants are seeking to restore the emotional equilibrium that the mother’s presence provided.
One of the Downstate separation experiments was physically as well as psychologically damaging to the monkey subjects. It was conducted over a decade ago but remains unpublished. A few details of the experiment were mentioned by Schlottmann and Seay (1972), who cited Kaufman’s data. Kaufman separated bonnet and pigtail infants and placed them with unfamiliar adult females. The infants “barely survived a single day” given the “brutality” of the females (Schlottmann & Seay 1972, p. 339).
University of Colorado Medical Center
Separation experiments have been conducted at the University of Colorado since the early 1970s. The subjects were pigtailed monkeys and, to a lesser extent, bonnet monkeys. The research was directed by psychiatrist Martin Reite, who focused on several physiological factors in the infant monkeys’ response to separation. These experiments were part of a trend begun in the early 1970s to add physiological data to the mass of behavioral data on infants’ responses to separation. These experiments were also viewed more generally as animal models of the increased susceptibility to disease and death following separation or loss of a loved one in humans. Reite claimed that the physiological responses to separation in monkey infants might throw light on the mechanism for this increased susceptibility in humans (Reite & Short 1983).
In the Colorado experiments, infant monkeys were housed with their mothers and other group members until they were five months old. At that time, mothers were removed for several days. The infants’ physiological patterns were monitored before, during and after separation via telemetry. A telemetry transmitter and power supply were surgically implanted in the infants’ abdominal walls, and electrode leads were “tunneled subcutaneously to their recording sites” (Reite & Short 1978, p. 1248).
The physiological responses to maternal separation included alterations in heart rate and rhythm, body temperature, sleep patterns, EEG patterns, and circadian rhythms. These results are summarized by Reite and Short (1983). The general conclusion from these studies is that maternal separation impairs physiological functions in infants by altering regulatory mechanisms.
The Colorado data on sleep disturbance add to the pathetic portrait of monkey infants whose mothers have been taken away. These infants take longer to fall asleep, awaken more frequently from sleep, and spend less total time asleep, than infants left with their mothers (Reite & Short 1978)
Reite et al. (1978) repeated some of their earlier experiments using cloth surrogate mothers instead of real mothers. For unexplained reasons, infants were reared with surrogates in social isolation, inside wooden crates. The physiological and behavioral responses to separation were less intense than those of infants reared with real mothers in social groups. The weak conclusion was that attachment bonds to cloth surrogates “may well be different than the bond to a living mother . . .” (p 427).
Reite et al. (1981a) also repeated some of their earlier studies, this time using a longer separation period (10 days). The longer separation was an attempt to observe the “recovery stage” noted in the Downstate experiments on pigtails. Little behavioral recovery occurred, however, providing another example of conflicting results from studies on the same species.
Two other studies focused on the apparent increase in disease and death following a separation or loss of a loved one in humans. One hypothesis for this increase is that separation or loss impairs the immune system. Two studies of human widows, cited by the Colorado team, strongly supported this hypothesis. Reite and coworkers tested and supported the same hypothesis. The experiments involved infant-infant separation (Reite et al. 1981 b) and mother-infant separation (Laudenslager et al. 1982).
In their most recent experiment, the Colorado researchers went to extraordinary lengths to induce depression in bonnet infants. Following Kaufman and Stynes (see above), Reite and Snyder (1982) separated a bonnet infant from its mother and all other bonnets in the group. This left the infant with unresponsive pigtails, and it became depressed. Its physiological responses to separation were similar to those seen in pigtails, notwithstanding the difference between separation from the mother (pigtails) and separation from all species members (bonnets).
The Kaufman and Stynes (1978) experiment discussed under Downstate experiments, for convenience, actually was carried out at Colorado, after Kaufman had moved from Downstate to Colorado.
Reite and Short (1983) recently found evidence that the physiological responses to maternal separation may result from factors other than the disruption of attachment bonds
. . . if a nursing infant [monkey] is deprived of the ability to nurse by putting a . . . vest on the mother, physiological changes similar to those seen during separation can be produced . . . (p. 247).
This experiment calls in question the interpretation of all the Colorado separation research. It also opens the door to a host of analytic experiments along the lines of Hofer’s rat studies.
Stanford University and Stanford Research Institute
Researchers at Stanford conducted several mother-infant separation experiments on primates since the late 1960s. Most of their subjects were squirrel monkeys. These monkeys inhabit forests in Central and South America; the adults used in the initial Stanford experiments had been purchased from an importer. A few Stanford separation experiments involved rhesus monkeys.
Most of these studies involved relatively brief (30-minute) separations and therefore focused on the panic that characterizes the immediate response to separation. Both hormonal and behavioral responses to separation were examined. The hormonal analyses were restricted to cortisol, which is associated with stress. Blood for these analyses was usually obtained by puncturing the heart of an anesthetized animal with a syringe. The same animals were sometimes subjected to this procedure more than once during a given experiment. When this happened, the monkeys were subjected to a maternal separation for each blood sample.
In the first Stanford separation experiment (Kaplan 1970), only behavioral responses to separation were investigated. Mother-infant pairs of squirrel monkeys were housed alone to preclude adoption of separated infants by other females. The infants were relatively independent of their mothers when separated and, not surprisingly, showed a relatively mild reaction to both separation and reunion. (One infant did die during separation, but apparently for unrelated reasons.) Kaplan attributed the mild reactions to the squirrel monkey’s permissive maternal style.
A cortisol analysis by Mendoza et al. (1978) indicated that a separation lasting 30 minutes was a “potent disturbance” to mother and infant squirrel monkeys. Rough handling (a part of the separation procedure) by itself elevated cortisol levels, but not if mother and infant were left together to comfort one another. The latter suggested that mother-infant interactions can suppress the hormonal response to stressful procedures.
This conclusion was also supported by a follow-up experiment involving the same separation procedure but a different blood sampling schedule (Levine et al. 1978). The opportunity for mutual comfort not only suppressed a cortisol response, but also reduced the cortisol response if the latter had already been activated.
In contrast to the Stanford studies summarized so far, most of the following studies involved mothers and infants living in groups, not in isolated pairs. For example, Coe et al. (1978) separated group-living mothers and infants to determine if adult females (“aunts”) could reduce the disturbance of separated infants. Disturbance levels were inferred from the infants’ hormonal and behavioral responses to separation. The behavioral response, but not the hormonal response, suggested that aunts could reduce the disturbance of separated infants. This decoupling of behavior and physiology suggested that behavioral quietude may mask shifts in internal state (Coe & Levine 1981). An earlier study also showed that aunts could reduce behavioral signs of disturbance in squirrel monkeys (Rosenblum 1971).
The Coe et al. study also suggested, along with a later study (Hennessy et al. 1979), that separation from a cloth surrogate mother is less disturbing than separation from a real mother. An earlier study (Mendoza et al. 1978) suggested that the response to separation from surrogates and real mothers was similar, but that study had weaker controls. Hence, the later experiments suggested that squirrel monkeys form greater attachment bonds to real mothers than to inanimate surrogates. Separation studies on rhesus monkeys and pigtailed macaques yielded similar results (see above).
Vogt et al. (1980) separated mother and infant squirrel monkeys and recorded the responses of nearby adults. Infants were removed from their group and caged either in or out of view of the adults. Only females showed an elevated cortisol response, with lactating females showing the highest response. The fact that non-lactating females responded at all was in sharp contrast with findings on rats . .” (p. 187). Among males and females, dominance ranks and cortisol levels were inversely related.
Vogt and Levine (1980) compared the responses of squirrel monkeys to partial separation (physical but not visual separation) and complete separation. The degree of separation did not affect the mothers’ or infants’ behavioral or hormonal responses. One reason offered for this result was that “. . brief separation seems so distressful for the squirrel monkey mother that the presence of her infant does not intensify her behavioral agitation” (p. 831).
This study raises several disturbing points regarding the value of this research. First, the results contrast with data from rhesus monkeys. The researchers noted that “there are several major differences between the present study and the previous research” on rhesus monkeys that confound comparisons (p. 831). One wonders how many studies would have to be done to reach a consensus of what the relationship is between degree of separation and response to separation. In the absence of a consensus, studies could be conducted indefinitely.
A second disturbing point is that cortisol levels are the lynchpin of this research, yet their interpretation is unclear. For example, an elevated cortisol level in one aspect of the present experiment may have resulted from “disturbance,” “fear,” “heightened arousal,” or “curiosity” (p. 831). Why bother measuring cortisol levels if their interpretation is so unclear? Marks (1981) was also skeptical of the Stanford researchers’ reliance on cortisol levels. In discussing these experiments, he noted:
. . . I am always a bit puzzled about the meaning of the cortisol response, especially in view of findings, for example, that comic films can produce as much increase in cortisol level in humans as can frightening films . . . (p. 175).
A third point concerns the degree to which the cortisol response system of squirrel monkeys is representative of primates. In regard to large differences between mothers’ and infants’ cortisol responses, the researchers wrote: “Other primate species apparently do not show these differences. . . . Under [baseline] and separation conditions rhesus mothers and infants exhibit comparable levels of plasma cortisol, as do human mothers and infants . ” (p. 832). The admitted aberrancy of the squirrel monkey’s cortisol response system calls into question the value of these separation studies.
In another Stanford study, Gunner et al. (1981) subjected squirrel monkey infants to a two-week separation. Separation induced brief cortisol elevations that returned to near-baseline levels within two hours. This suggested that the infants’ agitation was brief; however, some behavioral indications suggested that infants were agitated throughout the separation period. “Despair” responses were infrequently observed. Individual differences in cortisol responses to both separation and reunion were related to the degree of tension in the mother-infant relationship prior to separation. The researchers suggested that “for some infants, adapting to maternal loss may be less stressful than adapting to reunion with the mother” (p. 74).
A recent report summarized several new experiments (Coe et al. 1 983). In one experiment, mothers and infants were separated for 6 to 24 hours to provide data on a prolonged separation. Gunner et al. (see above) had already studied two-week separations, but apparently more data were deemed necessary. At any rate, the new study provided more evidence that behavioral and hormonal indicators of the protest response to separation could provide conflicting results.
A second experiment indicated that separated infants were more disturbed when housed in a strange environment than when left in the familiar environment. This experiment involved repeated four-hour separations; earlier experiments involving half-hour separations, but otherwise similar to the present experiment, were deemed too brief to assess the significance of strange vs. familiar separation environments. The present study agrees with a Cambridge study on rhesus monkeys (see above).
Another experiment summarized by Coe et al. examined the effects of repeated separations on the protest response. The infants’ behavioral responses (but not cortisol responses) attenuated over repeated separations. This result differed “radically” from Wisconsin data, but this discrepancy could have resulted from a multitude of factors, including the more benign procedures in the present experiment. The influence of procedural variation was inadvertently shown in a second experiment, which yielded increasing levels of behavioral response across separation.
The second study was a comparison of repeated separations with a fixed or variable duration. Fixed-interval separations elicited an increasingly severe behavioral disturbance, whereas variable-interval separations elicited a more stable and mild response. The researchers pointed out that these results are somewhat counterintuitive, because predictability is usually associated with less disturbance. However, the results are consistent with Levine’s hypothesis that in stressful situations, predictability without control over events may exacerbate, not ameliorate, the response.
Rhesus monkeys, not squirrel monkeys, were the subjects in a study by Smotherman et al. (1979). Both infants and mothers were removed from their group and then separated from one another (separation condition) or housed together (control condition). The same procedure on the same species was conducted at Cambridge but cortisol levels were not monitored in that study (see above). The cortisol levels of infants, but not of mothers, was higher in the separated condition. The rhesus mothers’ response contrasts with the response of squirrel monkey mothers summarized earlier. The infants’ cortisol response was related to maternal dominance rank. These conclusions were admittedly tentative owing to possible flaws in experimental design.
Miscellaneous Institutions
Many separation experiments were conducted at various institutions in addition to those mentioned above. Some of these studies involved the same primate species (rhesus, pigtailed, bonnet and squirrel monkeys) whereas other studies involved different species ( java monkey, paths monkey, langur, orangutan and gorilla).
Several studies added to the massive amount of data on rhesus monkeys accumulated at Wisconsin, Cambridge and Stanford. Three of these studies were conducted at the California Primate Center and involved Harlow’s former students, Mitchell and Mason.
- Brandt et al. (1972) attempted to compare infant responses to separation from the home cage and separation from the mother in the home cage. Infants in one group were reared alone and those in another group were reared with their mothers. The former were separated from their home cage, whereas the latter were left in the home cage but separated from their mother by a partition. Unfortunately, the home cages in the two groups were not the same; the isolates were reared in isolation chambers whereas the mother-reared infants were housed in wire cages. Given that the researchers were comparing apples to oranges, it is not surprising that they found several differences between the two groups in response to separation. Numerous conclusions were drawn from the experiment despite the authors’ recognition of this and other defects in experimental design.
- Several primate separation studies have shown that the presence of mothers reduces behavioral signs of insecurity in infants. Hill et al. (1973) showed the same effect on a physiological sign of insecurity. A high level of the hormone cortisol was the indicator of insecurity or stress. For unspecified reasons, the researchers used cloth surrogates instead of actual mothers. Moreover, all of these infants were reared in isolation, some in totally enclosed cages. A similar study with similar results was conducted at the Delta Primate Center (Candland & Mason 1969).
- Erwin (1974) observed the reaction of adult rhesus monkeys to recordings of a rhesus infant calling. The calls were taped immediately after the infant was separated from its mother. Females responded to the calls as if an infant were calling from their lap. Erwin did not use a neutral sound as a control, so he was forced to conclude that “there is no way of knowing from this study whether [the] subjects would have responded similarly to some other auditory stimulus . .” (p. 184).
Two other studies of rhesus monkeys were conducted at institutions other than the California Primate Center. In one of these studies, rhesus monkeys were subjected to a series of four mother-infant separations (Chappell & Meier 1975). Two involved separating infants from mothers (“infant-removed”) and two involved separating mothers from infants (“mother-removed”). The Cambridge researchers had already used the infant-removed versus mother-removed comparison, but they did not subject the same infants to both procedures. Chappell and Meier noted similarities and differences between their findings and those of the Cambridge studies. Also, it was noted that the repeated separations had diminishing effects, which contrasts with results from Suomi et al. (1970).
The other rhesus study extended separation experiments to the natural environment (Singh 1 975). Singh removed some mothers from two free-ranging groups in India and observed the response of their infants. The separated infants had to procure their own food during the 3-day separation period. The separated infants clearly were upset and restless. Singh mentions frequent “crying.” The results indicated that free-ranging rhesus infants became agitated by maternal separation. The separation period was too short to conclude that the same could be said for the despair phase.
Four separation studies of pigtailed monkeys were conducted at the University of Washington. These studies preceded the Colorado studies of pigtails, and emphasized mothers’ behavioral responses to separation, not the infants’ physiological responses. These studies are most note-worthy for their poignant descriptions of the monkeys’ behavior during the separation procedure and at reunion (see p. 78 above).
A separation study of squirrel monkeys was conducted at the University of Arizona (Jones & Clark 1973). These monkeys were living in groups, whereas the squirrel monkeys in the Stanford experiments that preceded this study were housed as solitary mother-infant pairs. Mothers were drugged to reduce the trauma of the separation procedure. The results were interpreted as consistent with the human protest-despair responses, although strong signs of despair (e.g. huddling) were not seen. Jones and Clark noted differences from the Stanford studies and offered post-hoc explanations for these differences.
Several new species were added to the list of primates involved in mother-infant separation studies. These species included patas monkey (Preston et al. 1970), java monkey (Schlottman & Seay 1972), Japanese monkey (Minami 1977), langur (Dolhinow 1980; Dolhinow & Murphy 1983), gorilla (Nadler & Green 1975), and orangutan (Nadler & Codner 1983). The experiments on patas and java monkeys were directed by Seay, a former student of Harlow. These studies provide further evidence of apparent species differences in response to separation.
Dolhinow studied langurs because infants receive substantial care from adult females other than their mothers, which is unusual among primates. When mothers of target infants were removed from the group, their infants showed signs of agitation followed by depression, but the response was extremely variable among infants. Two infants died. “Most infants adopted substitute caregivers in their mother’s absence, and a majority of infants elected to remain with [their] adopted female . . . when the mother returned to the group” (1983, p. 122). The two infants that died received too little substitute care. Dolhinow emphasized the importance of two variables in influencing a langur infant’s response to maternal separation: maternal restrictiveness and the availability of substitute caregivers. Those infants that reacted severely to maternal loss, including those that died, revealed that langur infants are strongly attached to their mother, despite contact with other adults.
Discussion
The Nature and Extent of Suffering
Separation experiments exact a heavy toll of animal suffering. Maternal separation causes acute distress in primates, dogs and cats. This distress gives way to despair in primates, and probably dogs and cats as well. Maternal separation can cause long-term psychological damage even if infants are reunited with their mothers. Long-term effects have been studied in primates; they include anxiety, anxious attachment to loved ones, and retarded social development. These effects are especially pronounced after repeated separations.
The damage from maternal separation experiments goes beyond psychological trauma. Physical damage, even death, was inflicted in scores of experiments. Rodents have borne the brunt of this treatment. They have been subjected to various surgical procedures and killed either during or after experiments. Primates suffered the same fate, but to a lesser extent. And primates, cats, and rodents have died from the separation experience alone.
Animals subjected to infant-infant separation, vertical chamber confinement, and cold-surrogate exposure also suffered greatly. Primates were the unwilling participants in these experiments. Those subjected to infant-infant separation seemed as seriously affected as infants separated from their mothers. Those subjected to vertical chamber confinement were psychologically crippled. Monkey infants reared with cold surrogate mothers were so devastated that some died.
Over 5,600 animals were subjected to these experiments. Rodents (4,847) and primates (579) were the workhorses of this research, with dogs (172) and cats (76) accounting for the balance. The last three figures are underestimates, as numerous studies did not report the numbers of animals used.
Financial Cost
Separation experiments cost over $34.0 million to conduct (Table IV). Most of this money was provided by the National Institutes of Health and the National Institute of Mental Health, as was the case with experiments in the deprivation category. Hence, U.S. taxpayers ultimately paid for most of this research.

Benefits to Humans
How have the results from separation experiments benefited humans? This is a fair question to ask given the clinical orientation of the majority of these experiments, not to mention the cost of the research to both animals and taxpayers.
Animal Model of Mother-Infant Separation
What have over two decades of animal experiments told us about mother-infant separation in humans? Judging from encouraging statements made by the animal modelers themselves (e.g. Brandt et al. 1972, p. 194; Harlow and Novak 1973, p. 464) one could easily assume that the animal studies have had considerable practical applications. But what does the evidence suggest?
Let us first consider the impact of the behavioral studies and then turn to the physiological studies. The behavioral studies apparently have failed to make any novel contribution to existing human data. The Wisconsin researchers themselves conceded this. After a strained attempt to establish the validity of their separation model, Suomi and Harlow (1977b) turned to the practical applications of the model: “We have a considerably more difficult time establishing a strong case [for practical applications], since so much monkey work to date has been based on existing human data and theories” (p. 173). Kraemer and McKinney (1979) reached the same conclusion: “it must be admitted that the primary flow of information, to date, has been from human clinical research into the animal modeling arena,” not vice versa (p. 38).
A comparison of animal and human studies suggests that, in general, the factors that influence the response to maternal separation are similar in human and nonhuman infants (Mineka & Suomi 1978; Mineka 1982). Although these animal studies did not make a novel contribution to the human situation, the rough correspondence between the animal and human data does lend support to the human data (Hinde 1974, Ainsworth 1976).
Animal models of the physiological responses to maternal separation have not fared any better than the behavioral models. A recent extensive review of the physiological experiments (Reite & Short 1983) did not mention a single clinical impact.
Separation studies of cats, dogs, and rodents seem to be even further removed from the human situation than studies of nonhuman primates. However, a few of these studies do make explicit reference to human applications. None of these applications seems particularly enlightening. For example, Elliot and Scott (1961) thought that their results on emotional distress in puppies suggested that “acute crying and fretting in human infants observed in day nurseries and hospitals may have more complex causes than simple separation from mother. Such infants are likewise on strange ground . . .” (p. 21). This patently obvious consideration appears in the early clinical literature (e.g. Yarrow 1961).
Some of the dog separation studies, like many of those on primates, appear to be reinventing the wheel; they go little beyond existing human data and theories. For example, Scott (1971) developed a theory of “attachment formation” on the basis of studies of dogs and humans. He wrote:
Bowlby and his colleagues [studying humans] have already done a great deal to develop ways of avoiding traumatic separation experiences in . . . infancy. The theory which I have developed here does not lead to any striking differences in procedures, but rather strengthens the reasons for them (p. 241).
Further evidence of the meager clinical impact that animal models of separation have had on our understanding of mother-infant separation in humans comes from several recent books on child development and early-childhood issues (Reference Note, p. 69). The books do not even mention any of these animal studies. Only human separation studies are discussed.
Animal models of mother-infant separation not only have failed to have any clinical impact, they also appear to have limited potential to make clinical contributions. The animal modelers have suggested two major aims for current separation studies (Hinde & McGinnis 1977, Dolhinow 1980, Reite & Short 1983): (1) identifying factors that increase susceptibility to the adverse effects of separation, and (2) discovering ways to ameliorate the adverse effects. Consider the procedures already known to ameliorate the response to maternal separation. First, human mothers have the ability to explain to the child the temporary nature and importance of the separation. Second, substitute care can be provided. That the effects of a separation experience in human children can be ameliorated or eliminated by sensitive fostering is well established” (Hinde & McGinnis 1977, p. 209). Third, mothers can attempt to meet the child’s increased emotional needs at reunion. Yarrow (1964) discusses numerous other ameliorating procedures.
Reite and Short (1983) speculate that animal models of mother-infant separation may be used for testing drugs to ameliorate the emotional distress of separated infants.
The evidence indicates that animal models of mother-infant separation in humans have had no clinical impact. At best, some of these experiments provided additional support of conclusions drawn from human studies. At worst, most of these experiments were trivial and redundant.
Separation Models of Human Depression
How have the prevention, diagnosis, treatment, and understanding of human depression benefited from animal studies of mother-infant separation, infant-infant separation, vertical chamber confinement, and other procedures?
One of the easiest ways to determine the clinical impact of animal models is to read the research reports of the animal modelers, who would not miss an opportunity to tout the importance of their work. None of the separation reports mentions any clinical impact of these studies.
Kraemer, Ebert, and McKinney (1983) suggested that their neurobiological studies of separation-induced despair have the potential to lead to clinical advances. They suggest that amine levels in cerebrospinal fluid could be used to identify individuals who are particularly vulnerable to separation-induced depression. These individuals could then be treated during separation to prevent depression. CSF amine levels may also be used to identify neurobiological targets for anti-depression drug therapy.
Time will tell whether or not these suggestions are correct; they don’t appear to be practical. It is difficult to imagine people seeking preventive therapy before they experience a disruptive separation. Moreover, CSF amine-directed drug therapy appears to be a red herring. Any of a number of pre-existing anti-depressants could be prescribed, if drug therapy is deemed necessary.
Separation Model of Anxiety
Suomi’s experiments on anxiety are too recent to have led to any clinical advances. Recall that his monkeys have temperamental differences that are apparent in stressful situations. These behavioral differences among individuals, and their associated physiological differences, are apparent early in development and are stable. In addition, they appear to be genetically based.
This research is highly regarded by the National Institute of Mental Health, which recently chose Suomi to head a new primate center (Cordes 1984). Moreover, this research apparently inspired parallel research on human infants that is currently underway (see Suomi 1986). What remains to be seen is whether or not these animal studies lead to clinical advances, and if so, whether or not these advances required animal studies.
Quality and Duplication
So far, separation experiments have a worse record on clinical application than do deprivation experiments. There is no suggestion that separation experiments on animals have had any clinical impact, while one deprivation experiment has had limited clinical impact.
The lack of clinical applications from separation studies would not be so alarming if few such studies were conducted. However, at least one hundred fifty scientific articles are devoted to these studies. Of these, about one hundred and ten correspond to separate studies. Most of these studies involved maternal separations in primates (Table III).
Why have so many experiments yielded so little?
One obvious reason is that the aim of many mother-infant separation experiments was to see if human responses, or factors that influence these responses, also occurred in this or that animal species. This was candidly acknowledged by Chappell and Meier (1975) who conducted a primate separation study themselves.
Another reason for the meager contribution of separation models is procedural variation from study to study. Studies differ in factors such as infants’ age at separation, degree of force used in the separation procedure, and degree of isolation imposed by the separation environment. These procedural variations can inadvertently affect the outcome of experiments. Moreover, if studies involve different animal species, then their outcomes can be affected by true species differences. While some of these variables are of interest in themselves, their effects are difficult to gauge when comparing studies that differ in more than one variable.
The problem of uncontrolled variation from study to study was the basis for Chappell and Meier’s (1975) pointed criticism:
Whereas these findings [from animal studies] are consistent, little resolution of the various issues central to maternal deprivation via separation can be gained until methodical variations are resolved. Variations in conditions in the supposedly controlled experimental work are as myriad as those in the naturally occurring experiments (p. 644).
Again:
The problems created by variations in design not only prevent comparisons across studies (only the grossest comparisons and conclusions can be drawn in this manner, i.e., that separation does result in agitation, at least, and depression if the period is long enough), but within a single study the findings are unclear (p. 645).
This criticism strikes at the heart of these experiments. Recall that the animal modelers originally exaggerated the inadequacies in human studies and touted the potential of their “well-controlled” experiments. Because of the unsystematic variation across studies, Mineka (1982) believes that the importance of variables such as infants’ age and sex has yet to be adequately investigated. This is a sad commentary given that scores of separation experiments have already been conducted.
What accounts for the lack of clinical applications of the various primate models of depression, namely, mother-infant separation, infant-infant separation, vertical chamber confinement, and cold surrogate exposure? One important factor is that virtually all of these experiments involve infants. Although separation models involving infants are thought to be scientifically valid models of human depression (Willner 1984), there are at least three reasons why infants are not good subjects for understanding human depression. One is that human depression is most prevalent in adults, not infants (Suomi & Harlow 1977b). Second, anaclitic depression (the despair reaction to maternal separation) in human children is not representative of adult depression (human data cited in McKinney 1977a and Suomi & Harlow 1977b). If this is true, then certainly anaclitic depression in monkeys is not representative of adult depression in humans. Third, an infant’s reaction to maternal separation is complicated by two factors (McKinney & Bunney 1969, Hofer 1978). One is the infant’s developmental progress, such as its increasing independence from its mother. The other complicating factor is the physiological as well as the psychological dependence of infants on their mothers. In fact, primate infants are so dependent on their mother that maternal separation can be a life-threatening event.
These shortcomings of using infants were apparent to the Wisconsin researchers themselves (Suomi & Harlow 1977b, McKinney & Bunney 1969, McKinney 1977a). Yet they justified their research at least partly—and in some cases wholly—as models of human depression. Consider Mineka’s (1982) justification: “depression in primates may best model childhood depression, which in turn may be thought of as a prototype for adult depression” (1982, p. 203). This view has two major problems. First, it ignores the human data that suggests anaclitic depression is not representative of adult depression. Second, it views the primate studies as models of models —twice removed from human adult depression.
The first of the maternal deprivation techniques to be explicitly viewed as a model of depression was maternal separation. However, the use of this model was challenged by Lewis et al. (1976) on the basis of unpromising results. “It becomes clear that mother-infant separation in rhesus monkeys does not fulfill the requirements of predictability and consistency, which are essential to establish a behaviorally-induced model of human depression” (p. 704, emphasis added). This criticism was not mentioned in Mineka’s (1982) extensive review of primate separation models of depression.
Lewis et al. advocated infant-infant separation as an alternative model of depression. However, they noted that researchers hesitated to use peer separation as a model of depression. According to Lewis et al., this hesitation might be related to the “absence of analogous human deprivations .” (Lewis et al. 1976, p. 705). If so, the hesitation seems justified. The use of peer separations violates one of McKinney and Bunney’s (1969) criteria for an animal model to be valid. Specifically, the procedure for inducing a syndrome in the model species should resemble the cause of the syndrome in humans. No one would argue that infant-infant separation, coupled with maternal deprivation, is a major cause of human depression.
Aside from maternal separation and peer separation, at least two other techniques were considered to be models of depression. One was confinement in vertical chambers and the other, exposure to cold surrogate mothers. Both were used exclusively at Wisconsin and were dismal failures.
The vertical chamber was initially heralded as having “enormous potential” as a model for human depression (Harlow & Suomi 1971b). Yet McKinney et al. (1975) later conceded that “there is considerable question as to what the behavioral changes induced by vertical chamber confinement represent” (p. 319). The vertical chamber was not even mentioned in Mineka’s (1982) extensive review of primate depression studies.
Mineka’s review also does not list exposure to cold surrogates as a model of depression. Yet this lethal technique was initially heralded as a model of “extreme depression” (Harlow et al. 1973). Harlow and company apparently were the only researchers to consider it as such. Actually, Harlow originally thought this procedure would be an animal model of schizophrenia (Harlow, in Tavris 1973), but that also was wishful thinking.
The Wisconsin researchers applied clinical labels so carelessly to their animal models that even isolation chamber confinement was eventually labeled a model of depression (Harlow 1974). Alas, this was also wishful thinking.
One of the most unsettling criticisms of primate models of depression is that depression in monkeys lacks salient cognitive features of depression in humans. These cognitive features include negative thoughts about oneself, the world, and the future.
Many researchers have questioned the usefulness of primate models of depression partly because, they argue, many of the cognitive symptoms which have been claimed to be cardinal symptoms in humans may either not exist in primates, or at a minimum may be very difficult to model in primates (Mineka 1982, p. 201).
Mineka, a Wisconsin researcher, gets around this criticism by arguing that these cognitive features are not cardinal symptoms of depression, and therefore are not needed to diagnose depression.
Researchers have other grounds for doubting that separation in monkeys is even a model of human depression, let alone a good model (e.g. Engel & Schmale, cited in Kaufmann 1977). Ainsworth (1976), for example, took issue with labeling the despair following separation as “depression.” She observed that the use of this label “implies that [the despair response] is akin to depressive illness in adults.. . . There is nothing essentially pathological in grief and mourning” following loss of a loved one (p. 41). This potentially lethal criticism remains unanswered. It and other criticisms go unheeded as researchers continue to tout the applicability of their work to human depression.
Alternatives
Most of the animal separation studies concerned the response to separation and the factors that influenced this response. The irony in this is that the “alternative,” human studies, preceded the animal studies, as was the case with studies of maternal deprivation per se (Ch. 4).
Not all separation studies were attempted replications of human research. Two noteworthy examples are the recent studies by McKinney’s group and Suomi’s group. If both lines of research prove to be valuable, their value will be in engendering parallel human studies. Hence, the animal studies would be facilitative, not essential.
Hinde’s group at Cambridge showed that primate separation studies could be conclusive without being excessively restrictive. Monkey infants could be housed with species members in large outdoor enclosures, not in tiny cages with or without their mother. Monkey infants did not have to be confined in vertical chambers to show the importance of the separation environment. They did not have to be separated up to 28 times to show the effects of more than one separation.
Summary
Separation experiments involved separating infants from their mothers or peers after the infants had developed attachment bonds. Related procedures involved incarcerating infants in vertical chambers or exposing them to cold surrogate mothers. These studies were animal models of mother-infant separation, depression, and anxiety.
Over 5,600 animals, mostly primates and rodents, were subjected to these experiments. The primates suffered acute distress and despair, with signs of chronic anxiety. Some were killed. The rodents suffered more physical damage; many were killed.
These experiments cost over $34 million, supplied primarily by federal institutions.
Although over 100 separation experiments have been conducted, none has yet had an impact on clinical practice. Many experiments were trivial extensions of past research, including human research, or were misconceived or misdesigned. Moreover, alternatives to many of these experiments were either already in use or could have been devised.
.org/maternal-deprivation-experiments-psychology/chapter-six/”>style=”color: #808080;”>
Please cite as:
Stephens, Martin L. (1986) Maternal Deprivation Experiments in Psychology: A Critique of Animal Models.
Retrieved from http://aavs.org/maternal-deprivation-experiments-psychology.
General Discussion
Over 7,000 animals, including primates, dogs, cats and rodents, were subjected to maternal deprivation experiments (Table V). The deprivation experience was often coupled with stressful tests or surgery. The deprivation experience alone was enough to cause psychological damage, which ranged from acute distress to generalized psychological crippling. Many animals succumbed to the experimental procedures or were killed outright.

The price tag of these experiments was over $57 million (Table VI). Virtually all of this money came from agencies of the federal government, such as the National Institutes of Health and the National Institute of Mental Health. This indicates that taxpayers have unwittingly financed most of these experiments.
The search for direct clinical impacts of these animal experiments has nearly been in vain. Only one experiment had had a direct clinical impact. It involved habilitating monkeys reared in isolation. The habilitation procedure entailed pairing these disturbed monkeys with younger, more-normal individuals. This procedure has been applied successfully to shy children and children at risk for schizophrenia.
There is a double irony in this application of animal data. Not only did the habilitation procedure involve well-known psychiatric principles, it also could have been safely developed in human studies, without prior animal studies. Moreover, in the case of the shy children, the reason for the procedure’s success was the reverse of that in the animal study.
Other experiments have had little or no clinical impact. At best, the scores of deprivation and separation experiments simply added support to pre-existing human data. Only a handful of experiments went beyond the human data to make novel suggestions about clinical problems.
To date, several lines of physiological research on maternal deprivation or separation have failed to have any clinical impact. Except for McKinney’s research, much of this work appears to be basic research far removed from clinical concerns.

Hence, despite the intentions of the researchers, their experiments have made only a small contribution to clinical practice. This is a serious charge. The animal modelers could attempt to offset this by highlighting the theoretical implications of their research. Animal modelers could present at least two other defenses of their maternal deprivation experiments. The first is that the potential value of these experiments has been largely overlooked by clinicians, and, that, in time, this value will be appreciated. The second is that more experiments are needed before human benefits can be realized.
All three of these arguments have been made in a general defense of psychological experiments on animals (Gallup & Suarez 1980). However, skeptics would regard these defenses as excuses or vacuous enticements to lure further funding. A larger measure of skepticism is warranted. None of the reports cited in this survey provides examples of theoretical implications that have led to human research with practical application.
Three decades of maternal deprivation experiments on animals have produced over 350 research reports. Not all of these reports constitute separate studies; some discuss previously published experiments. However, there are at least 250 separate studies. This figure does not include unpublished experiments that are completed or underway. Some of these experiments will never be published.
Why were so many maternal deprivation experiments conducted? The seminal finding of separation experiments was that rhesus monkeys exhibit, under certain conditions, a protest-despair response similar to humans. Subsequent experiments were attempts to demonstrate the same response in other species or in rhesus monkeys reared in different conditions. These types of studies are still being conducted.
Other experiments were attempts to determine the influences of several variables on an infant’s behavioral responses to separation. These variables included the infant’s age, length of separation, degree of separation, previous history of separation, nature of mother-infant relationship, nature of separation environment, and other factors. The effect of these variables on a variety of physiological responses to separation was also studied. Different species, as well as different responses, were studied.
Mother-infant separation experiments led the Wisconsin researchers into their depression research. Suomi began his peer separation experiments to find a new way of inducing depression. The same sorts of experiments that involved mother-infant separation could be conducted again using infant-infant separations. Harlow introduced his vertical chamber as another means of inducing depression.
Mother-infant separation experiments also led the Wisconsin researchers into their recent anxiety research.
The clear message of this scenario is that these studies could drag on indefinitely, with yet more species, more diverse situations, and more procedures being tested. Indeed, recent reviews of separation experiments state that many more experiments need to be conducted. One is reminded of Levy’s (1952) adage: The “animal psychologist . . . is often wedded forever to his animal or his instrument” (p. 485).
The point is not that all maternal deprivation experiments were exact duplicates of others, or that none of these experiments had scientific merit. The point is that most seem trivial in light of their harsh exploitation of animals, their dismal record on clinical applications, their mediocre quality, and the value of alternative, human studies.
Consider the problem of research quality. Several lines of research used poor models of human phenomena. Total isolation was not a good model of institutionalization; separation of infant monkeys from their mothers when the latter were the infants’ only companion was not a good model of the more complex human situation; vertical chamber confinement was not a good model of depression, etc. Some poor models were quickly abandoned but others were used again and again. For example, infant monkeys were subjected to just about every conceivable schedule of total isolation.
The quality of maternal deprivation research was compromised a priori when poor models were used. Research quality was further compromised by problems with experimental design. Numerous experiments had defective designs. Moreover, the designs of separation experiments were nonstandardized, so that results differing from study to study could have resulted from any of a number of factors.
The problem of self-perpetuation of experiments, at least insofar as maternal deprivation experiments are concerned, is not one of inadequate dissemination of scientific information. Researchers propose their new, slightly different experiments, not out of ignorance of previous research, but in an effort to add to previous research (however trivial the additional information). In some ways, past research encourages new research, producing a bandwagon effect. Hence, this survey supports Gendin’s (1984) belief that passing legislation to better disseminate scientific information will not necessarily reduce duplicative research, but might even encourage it.
The topic of alternatives is almost moot with respect to models of maternal deprivation and maternal separation. After all, human studies predated these animal models. One wonders what conclusions the modelers would have drawn without the human data for direction and comparison. Recall that animal species differed in their basic behavioral response to deprivation and separation, not to mention responses to subtler variations in procedures.
The maternal deprivation modelers have overly criticized human studies. To be sure, the human research contained numerous examples of poor design, and it relied substantially on retrospective studies. Examples of faulty research can be found in any field. However, the animal modelers were attempting to throw the baby out with the bathwater. The human research contained numerous examples of well-designed and/or prospective studies (see Yarrow 1961). With all their purported drawbacks, human studies of maternal deprivation and maternal separation were the first to yield the conclusions that the animal studies later “supported.”
Research strategies for studying maternal deprivation and separation in human infants often involve “natural experiments.” Yarrow (1968) discusses two such situations:
[1] Child-care institutions that differ in basic child-rearing philosophies. There are great variations among these institutions in caretaker ratios and in various characteristics of the physical-sensory environment, such as . . . complexity of visual and auditory stimulation. Even though past studies of these settings have been theoretically unsophisticated and have had serious methodological shortcomings, they have produced some significant findings. With sharper conceptualizations and more refined observational techniques, more precise data on the effects of very early sensory and social deprivation in human infants can be obtained. Moreover, such settings also lend themselves to simple enrichment studies involving experimental intervention, e.g., . . . training caretakers to give measured degrees of language or social stimulation, or to respond contingently to specific aspects of the infant’s behavior.
[2] Several kinds of separation situations that occur with moderately great frequency in the life of young children allow exploration of important aspects of early experiences. Among the many kinds of situations in which variables such as loss of a significant relationship figure, broad environmental changes, or novelty, can be teased out and studied independently or in interaction are the following. A) Situations in which the mother leaves the infant temporarily in familiar surroundings, as distinguished from temporary separation associated with gross environment change. B) Situations in which the infant is temporarily moved to another home with an attentive caretaker, as distinguished from separation followed by gross deprivation of maternal care, as in institutionalization. C) Separation experiences associated with adoptive placement involving a change in mother figure (p. 107-108).
Yarrow adds:
The use of natural situations in research has inherent difficulties because of the many variables in complex and uncontrolled interactions, only some of which we are aware of and can specify. To use natural situations effectively, it is necessary that we conceptualize as clearly as possible the theoretical issues, and identify clearly the variables in the situation relevant to the theoretical issues , . . (p. 108).
Observational techniques for such human studies have become increasingly sophisticated over the years. Ainsworth (1976) has incorporated ethological techniques into her naturalistic studies of mothers and infants. Unfortunately, “naturalistic studies of [human] mother-infant interactions . . . are not popular with grant-awarding panels” (p. 46).
Naturalistic studies examine a process in the context of its ecological setting. In contrast, experimental studies with animals are often attempts to remove influences of all variables except one.
The problem with this approach is that in order to keep all other factors constant, infants must be reared in relatively sterile environments that do not promote normal social development (Suomi 1977b, p. 212).
Yarrow (1968) highlighted the importance of naturalistic studies in unraveling human development.
The values and limitations of experimental studies with animals under closely controlled conditions should be weighed against the values and limitations of investigations with humans in naturalistic settings. Some kinds of problems clearly lend themselves to experimental manipulation of a limited number of variables; for other kinds of problems, such an approach can yield only suggestive hypotheses that must be tested in more complex settings. There are obvious limitations in the extent to which relationships established in a controlled “pure situation” can be generalized to the complex naturalistic setting. The action of a single variable in a hypothetical vacuum may be very different from the action of this same variable in a network of other variables. In environments in which there are many variables in complex interaction, as in most human environments, the significance of any given variable or limited set of variables may be radically altered by the larger context. . . As the behavioral sciences become more mature, I think we will have to give up the inordinate value placed on experimental manipulation of a few simple variables, and give increasing attention to the development of research designs for controlled analyses of complex environments (p. 112-13).
Greater understanding of mother-infant interactions and child development can also be achieved through increased use of cross-cultural human studies. Although this kind of research has inherent weaknesses, these seem no greater than those handicapping animal studies. Cross-cultural studies can also reveal that conclusions from animal data are culture-bound. What animal data tell us about “the human case” depends on what we regard as the human case. With respect to the monkey separation studies, “what we have been pleased to call the human case” is Western society (Lehrmann 1974, p. 194).
Animal Models in Psychiatry
Most of the animal models discussed above can be considered to fall within the domain of psychiatry. These include models of mental illness, maternal deprivation, maternal separation, and depression. These models were part of a move toward using animal models of psychiatric disorders. This move was prominent during the 1960s and was an attempt to mimic animal model research in other fields of medicine (McKinney et al. 1975, Reite 1977).
Psychiatrists have traditionally been skeptical of animal models of mental disorders. This skepticism stems partly from the belief that human mental capacities are necessary to become neurotic (McKinney 1974a), and was fueled by careless analogies in early animal research (Reite 1977). This skepticism among psychiatrists still lingers. For example:
Human nature can not be reduced to experimental models of laboratory animals. . . . lf, however, any scientific conclusions are to be drawn from the animal model, the animal should be observed in the natural habitat . . . (Serban 1976, p. 280).
Again, “mechanical concepts [of experimental psychologists] are totally unsuited to man’s psychosocial existence” (Serban, in Serban et al. 1976, p. 2).
However, according to Suomi (1984a), “animal models of human psychopathology have been gaining increasing respectability and use” (p. 226). Several of the maternal deprivation modelers themselves have been psychiatrists, including McKinney, Reite, Kaufman, Levine, Jensen and Hofer.
Ironically, as the psychiatrists’ skepticism of animal models is diminishing, the animal modelers themselves are beginning to acknowledge that animal models at best can make only tentative suggestions about human parallels (e.g. Suomi 1982). This implies that poor animal models yield suggestions about humans that are indeed shaky. Consider a recent separation study of rhesus monkeys by Suomi et al. (1983). The researchers wrote:
What relevance, if any, do these findings have for . . . human mothers (and fathers) and infants? We clearly acknowledge that cross-species generalizations should be made with great caution and according to well-thought-out principles and that true one-to-one matches of phenomena between species are relatively rare. . . . Furthermore, the living conditions of our monkeys during separations are not analogous to the living conditions of most human infants during parental separations and it seems unlikely that exactly the same pattern of results would occur if our infant monkeys had been with peers or other adults during their separations, as human infants usually are. Nevertheless, the present monkey data raise some issues that might profitably be kept in mind when pondering possible consequences of frequent parental separations in young children (p. 784).
Other statements in the maternal deprivation literature also do not inspire confidence in animal models. For example, Levine (in Harlow & Suomi 1976) once pointed out to Harlow that animals in laboratories are abnormal, even before experimental manipulations. Harlow’s response included the following statement: “You pray that [the research measures] have validity; . . . you just have to gamble!” (p. 74).
In spite of the shortcomings of animal models, the Wisconsin researchers have vigorously promoted their use in psychiatry.
The major areas of interest for psychiatrists in primate models probably include the development of kinds of attachment behavior, the effects of social isolation, separation studies, the possible experimental simulation of learned helplessness, and various biological approaches being developed. It is in these areas that work has been and is being conducted that has potential clinical usefulness. Investigators may be close to developing a viable animal model for depression that may facilitate a more comprehensive understanding of this particular syndrome and enable studies to be done that are currently impossible to perform utilizing human beings. The rest of medicine has long used nonhuman primates to advance knowledge about their fields and there is no logical reason why psychiatry should not do the same (McKinney et al. 1975, p. 330).
Yet, skepticism of animal models should remain firm. At least three reasons are apparent. First, experiments have had very little clinical impact. Second, they siphon money away from acceptable research on the human condition. Third, they subject animals to harsh treatment.
Maternal deprivation researchers have almost completely avoided discussing humane and ethical issues raised by their callous treatment of animals. There is virtually no indication that any procedures were designed or modified out of compassion for the animal subjects. The use of anesthesia and similar procedures was based on efficacy, not humanity.
Maternal deprivation researchers raise ethical issues only when “explaining” that ethical constraints dictate that animals be used instead of humans. The implicit premise is that the experiments can be performed on animals because animals are different from humans. The irony is that the scientific rationale for these experiments emphasizes the similarities between animals and humans. This similarity is demonstrated by the experiments themselves. A few researchers were struck by this similarity (e.g. Suomi 1977b, p. 222). Consider the conclusion from Bronfenbrenner’s comparison of the effects of deprivation-rearing in human and nonhuman primates:
. . . the most surprising and possibly sobering outcome of [our] . . . analysis [is] the similarity we humans bear to our fellow primates — at least when it comes to our need for stimulation and for attachment to others (1968, p. 756).
Unfortunately, mounting evidence of the similarities between human and nonhuman primates has not engendered a reevaluation of maternal deprivation experiments on animals. Instead, the similarities are considered an incentive for further research.
This situation puts the animal modelers in an odd position. On the one hand, their experiments raise disturbing questions about the propriety of this research. Moreover, public concern over these experiments is mounting. On the other hand, the modelers want to continue their research. The same dilemma applies to animal research in general. Researchers and funding agencies have responded by proposing guidelines for animal research. However, as in many cases of self-regulation, the strength of these guidelines has been exaggerated. Consider Suomi’s (1984b) statement:
. . . in most respects, current scientific and ethical guidelines for care and treatment of nonhuman primate subjects are at least as strict as those pertaining to research utilizing human subjects (p. 120).
Suomi does grant that the privilege to use primates in research does not mean that researchers have free reign over what they can do to their subjects.
What it does mean is that experimenters can selectively breed specific rhesus monkey pairs, and they can perform cross-fostering manipulations in order to separate possible genetic and rearing environ-mental effects. Moreover, they can monitor their monkey subjects’ behavioral and physiological activities to a degree and duration almost never feasible for any humans . . . (Suomi 1984b, p. 120).
Suomi neglected to mention harsher procedures that apparently are still allowed under the research guidelines. These include traditional deprivation experiments (e.g. Coelho & Bramblett 1984) and separation experiments (e.g. Nadler & Codner 1983).
The National Institute of Mental Health (NIMH) is worried about the public response to psychology experiments on animals. After NIMH recently hired Suomi, its director of intramural research praised him as “an animal protectionist of the first class” for his concern about the care of research animals (Cordes 1984). This concern for animals is not revealed in Suomi’s research reports. Consider the following passage, which was a discussion of his research plans:
We are also interested in the tunnel of terror in that it may be utilized in various ways in combination with the pits [vertical chambers], to shorten, intensify, and possibly stereotype the depressive syndrome. For example, individual monkeys or social pairs may be driven to the pits by the gradual or rapid approach of the marching monster. Depression in individual monkeys already incarcerated may be further enhanced by the appearance or advance of the monster from the top or ends of the pit chambers. Also, pairs of monkeys may be separated after tunnel terror or monster machinations in order to assess the possible complementary contribution of social separation to psychopathological production (Suomi & Harlow 1969, p. 249).
Characterizing people who conduct this type of research as animal protectionists does not instill confidence in the self-regulation of psychological research on animals.
Summary and Conclusions
Over 250 maternal deprivation experiments on animals have been conducted to date. These experiments cost over $57 million, provided primarily by the federal government. Over 7,000 animals were subjected to procedures that induced distress, despair, anxiety, general psychological devastation, or even death. The results of this research have had little impact on clinical practice, and the potential for future advances seems limited. Many experiments were trivial extensions of past research, or simply were attempts to reproduce in animals what was already known about humans.
Given research such as this, it is not surprising that psychology experiments on animals have been criticized as cruel, wasteful or misguided (e.g. Bowd 1980, Bannister 1981, Drewett & Kani 1981, McArdle 1984). Rebuttals of these charges (Gallup & Suarez 1980; Coile & Miller 1984, Miller 1985) illustrate the need for in-depth evaluations of psychological research. The present evaluation provides substantial support for the critics, at least with respect to maternal deprivation experiments. If this area of research is representative of others within psychology, then the use of animals in psychological research faces a formidable challenge.
Please cite as:
Stephens, Martin L. (1986) Maternal Deprivation Experiments in Psychology: A Critique of Animal Models.
Retrieved from http://aavs.org/maternal-deprivation-experiments-psychology.
Ackerman, S. H., Hofer, M. A. & Weiner, H. 1975. Age at maternal separation and gastric erosion susceptibility in the rat. Psychosom. Med. 37: 180-184.
Ackerman, S., Hofer, M. A. & Weiner, H. 1978. Early maternal separation increases gastric ulcer risk in rats by producing a latent thermoregulatory disturbance. Science 201: 373-376.
Ader, R. 1962. Social factors affecting emotionality and resistance to disease in animals: III. Early weaning and susceptibility to gastric ulcers in the rat: a control for nutritional factors. J. Comp. Physiol. Psychol. 55: 600-602.
Ader, R. & Friedman, S. B. 1965. Social factors affecting emotionality and resistance to disease in animals: V. Early separation from the mother and response to a transplanted tumor in the rat. Psychosom. Med. 27: 119-122.
Ader, R., Tatum, R. & Beels, C. C. 1960. Social factors affecting emotionality and resistance to disease in animals: I. Age of separation from the mother and susceptibility to gastric ulcers in the rat. J. Comp. Physiol. Psychol. 53: 446-454.
Agrawal, H. C., Fox, M. W & Himwich, W. A. 1967. Neurochemical and behavioral effects of isolation-rearing in the dog. Life Sci. 6: 71-78.
Ainsworth, L. L. 1984. Contact comfort-A reconsideration of the original work. Psychol. Rep. 55: 943-949.
Ainsworth, L. L. & Baker, C. D. 1982. Effects of illumination changes on infant monkey contacts with surrogates. Psychol. Rec. 32: 513-518.
Ainsworth, M. D. 1962. The effects of maternal deprivation: a review of findings and controversy in the context of research strategy. Pp. 97-165 in: Deprivation of maternal care: a reassessment of its effects. Geneva: WHO Public Health Papers, no. 14.
Ainsworth, M. D. S. 1976. Discussion of papers by Suomi and Bowlby. Pp. 37-47 in: Animal models in human psychobiology (G. Serban & A. Kling, eds.). New York: Plenum.
Ambron, S. 1984. Child development. New York: Holt, Rhinehart & Wilson.
Anderson, C. O., Kenney, A. M. & Mason, W. A. 1977. Effects of maternal mobility, partner, and endocrine state on social responsiveness of adolescent rhesus monkeys. Dev. Psychobiol. 10: 421-434.
Anderson, C. O. & Mason, W. A. 1974. Early experience and complexity of social organization in groups of young rhesus monkeys (Macaca mulatta). J. Comp. Physiol. Psychol. 87: 681-90.
Andry, R. G. 1962. Paternal and maternal roles and delinquency. Pp. 31-44 in: Deprivation of maternal care: a reassessment of its affects. Geneva: WHO Public Health Papers, no. 14.
Angermeier, W F., Phelps, J. B., Murray, S & Reynolds, H. H. 1967. Dominance in monkeys: early rearing and home environment. Psychon. Sci. 9: 433-434.
Arling, G. L. & Harlow, H. F. 1967. Effects of social deprivation on maternal behavior of rhesus monkeys. J. Comp. Physiol. Psychol. 64: 371-378.
Arling, G. L., Ruppenthal, G. C. & Mitchell, G. D. 1969. Aggressive behaviour of the eight-year-old nulliparous isolate female monkey. Animal Behay. 17: 109-13.
Bannister, D. 1981. The fallacy of animal experimentation in psychology. Pp. 307-317 in: Animals in research (D. Sperlinger, ed.). New York: John Wiley & Sons, Ltd.
Baysinger, C., Brandt, E. M. & Mitchell, G. 1972. Development of infant social isolate monkeys (Macaca mulatta) in their isolation environments. Primates 13: 257-270.
Baysinger, C. M., Plubell, P E. & Harlow, H. F. 1973. A variable – temperature surrogate mother for studying attachment in infant monkeys. Behav. Meth. and Instr. 5: 269-272.
Beach, F. A. & Jaynes, J. 1954. Effects of early experience upon the behavior of animals. Psychol. Bull. 51: 239-263.
Beasley, J. W. & Seal, U. S. 1968. Effects of age and rearing conditions of Macaca mulatta on blood proteins and protein-bound carbohydrates. Clin. Chem. 14: 1107-1111.
Beckett, P. G. S., Frohman, C. E., Davenport, R. K. G., Jr., Rogers, C. M. & Gottlieb, J. S. 1963b. Biologic effects of infantile restriction in chimpanzees. Compr. Psychiat. 4: 1-8.
Beckett, P. G. S., Frohman, C. E., Gottlieb, J. S., Mowbray, J. B. & Wolf, R. C. 1963a. Schizophrenic-like mechanisms in monkeys. Am. J. Psychiat. 119: 835-842.
Berkson, G. 1967. Abnormal stereotyped motor acts. Pp. 76-94 in: Comparative psychopathology (J. Zubin & H. F. Hunt, eds.). NY: Grune & Stratton.
Berkson, G. 1968. Development of abnormal stereotyped behaviors. Dev. Psychobiol. 1: 118-132.
Berkson, G., Goodrich, J. & Kraft, 1. 1966. Abnormal stereotyped movements of marmosets. Perc. Motor Skills 23: 491-498.
Berkson, G. & Mason, W. A. 1964. Stereotyped behaviors of chimpanzees: Relation to general arousal and alternative activities. Perc. Motor Skills 19: 635-652.
Berkson, G., Mason, W. A. & Saxon, S. V. 1963. Situation and stimulus effects on stereotyped behaviors of chimpanzees. J. Comp. Physiol. Psychol. 56: 786-792.
Bowd, A. D. 1980. Ethical reservations about psychological research with animals. Psychol. Rec. 30: 201-210.
Bowlby, J. 1952. Maternal care and mental health. Geneva: WHO Monograph Series, No. 2.
Bowlby, J. 1958. The nature of the child’s tie to his mother. Int. J. Psycho-Anal. 39: 350-373.
Bowlby, J. 1976. Human personality development in an ethological light. Pp. 27-36 in: Animal models in human psychobiology (G. Serban & A. Kling, eds.). New York: Plenum
Brandt, E. M., Baysinger, C. & Mitchell, G. 1972. Separation from rearing environment in mother-reared and isolation-reared rhesus monkeys (Macaca mulatto) Int. J. of Psychobiol. 2: 193-204.
Brandt, E. M. & Mitchell, G. 1973. Pairing preadolescents with infants (Macaca mulatta) Dev. Psychol. 8: 222-228.
Brandt, E. M., Stevens, C. W. & Mitchell, G. 1971. Visual social communication in adult male isolate-reared monkeys (Macaca mulatta). Primates 12: 105-112.
Breese, G. R., Smith, R. D., Mueller, R. A., Howard, J. L., Prange, A. J., Jr., Lipton, M. A., Young, L. D., McKinney, W. T. & Lewis, J. K. 1973. Induction of adrenal catecholamine synthesising enzymes following mother-infant separation. Nature- New Biology 246: 94-96.
Bronfenbrenner, U. 1968. Early deprivation in mammals: a cross-species analysis. Pp. 627-764 in: Early experience and behavior (G. Newton & S. Levine, eds.). Springfield, Ill.: Charles Thomas.
Candland, D. K. & Mason, W. A. 1969. Infant monkey heart rate: habituation and effects of social substitutes. Dev. Psychobiol. 1: 254-256.
Capitanio, J. P. 1984. Early experience and social processes in rhesus macaques (Macaca mulatta) I. Dyadic social interaction. J. Comp. Psychol. 98: 35-44.
Casler, L. 1961. Maternal deprivation: a critical review of the literature. Monogs. Soc. Res. Child Develop. 26, No. 2.
Chalmers, N. R. & Locke-Haydon, J. 1984. A comparison of the effects of short-term separation from caregivers and of caregiver indifference on infant behavioural development in captive marmosets, Callithrix jacchus jacchus. Int. J. Primatol. 5: 327 (abs.)
Chamove, A. S. 1973a. Rearing infant rhesus together. Behaviour 47: 48-66.
Chamove, A. S. 1973b. Social behavior comparison in laboratory reared stumptail and rhesus macaques. Folia Primatol. 19: 35-40.
Chamove, A. S. 1978. Therapy of isolate rhesus: Different partners and social behavior. Child Devel. 49: 43-50.
Chamove, A. S., Rosenblum, L. A. & Harlow, H. F. 1973. Monkeys (Macaca mulatta) raised only with peers: a pilot study. Anim. Behay. 21: 316-325.
Chappell, P. F. & Meier, G. W. 1975. Modification of the responses to separation in the infant rhesus macaque through manipulation of the environment. Biol. Psychiat. 10: 643-657.
Codner, M. A. & Nadler, R. D. 1984. Mother-infant separation and reunion in the great apes. Primates 25: 204-217.
Coe, C. L. & Levine, S. 1981. Normal responses to mother-infant separation in nonhuman primates. Pp. 155-175 in: Anxiety: New research and changing concepts (D. F. Klein & J. G. Rabkin, eds.). New York: Raven.
Coe, C. L., Mendoza, S. P., Smotherman, W. P. & Levine, S. 1978. Mother-infant attachment in the squirrel monkey: adrenal response to separation. Behay. Biol. 22: 256-263.
Coe, C. L., Wiener, N. & Levine, S. 1983. Psychoendocrine responses of mother and infant monkeys in disturbance and separation. Pp. 189-214 in: Symbiosis in parent-young interactions (H. Moltz & L. A. Rosenblum, eds.). New York: Plenum.
Coelho, A. M., Jr. & Bramblett, C. A. 1981. Effects of rearing on aggression and subordination in Paplo monkeys. Am. J. Primatol. 1: 401-412.
Coelho, A. M., Jr. & Bramblett, C. A. 1984. Early rearing experiences and the performance of affinitive and approach behavior in infant and juvenile baboons. Primates 25: 218-224.
Coile, D. C. & Miller, N. E. 1984. How radical animal activists try to mislead humane people. Am. Psychol. 39: 700-701.
Comroe, J. H., Jr. & Dripps, R. D. 1976. Scientific basis for the support of biomedical science. Science 192: 105-111.
Cordes, C. 1984. Suomi heads new primate center. Am. Psychol. Assoc. Monitor, August, 10.
Cross, H. A. & Harlow, H. F. 1965. Prolonged and progressive effects of partial isolation on the behavior of macaque monkeys. J. Exp. Res. in Pers. 1: 39-49.
Cullen, J. H., ed. 1974. Experimental behaviour: a basis for the study of mental disturbance. New York: J. Wiley & Sons.
Cummins, M. S. & Suomi, S. J. 1976. Long-term effects of social rehabilitation in rhesus monkeys. Primates 17: 43-51.
Damon, W. 1983. Social and personality development: Infancy through adolescence. New York: Norton.
Davenport, R. K., Jr. & Menzel, E. W., Jr. 1963. Stereotyped behavior of the infant chimpanzee. Arch. Gen. Psychiat. 8: 99-104.
Davenport, R. K., Jr. Menzel, E. W., Jr. & Rogers, C. M. 1961. Maternal care during infancy: its effect on weight gain and mortality in the chimpanzee. Amer. J. Orthopsychiat. 31: 803-809.
Davenport, R. K., Menzel, E. W, Jr. & Rogers, C. M. 1966. Effect of severe isolation in “normal” juvenile chimpanzees. Arch. Gen Psychiat. 14: 134-138.
Davenport, R. K., Jr. & Rogers, C. M. 1968. Intellectual performance of differently reared chimpanzees: 1. Delayed response. Am. J. Ment. Defic. 72: 674-680.
Davenport, R. K. & Rogers, C. M. 1970. Differential rearing of the chimpanzee. A project survey. Pp. 337-360 in: The chimpanzee, Vol. 3 (G. H. Bourne, ed.). Basel: S. Karger.
Davenport, R. K., Jr., Rogers, C. M. & Menzel, E. W., Jr. 1969. Intellectual performance of differentially reared chimpanzees: II Discrimination-learning set. Am. J. Ment. Defic. 73: 963-969.
Davenport, R. K., Rogers, C. M. & Rumbaugh, D. M. 1973. Long-term cognitive deficits in chimpanzees associated with early impoverished rearing. Devel. Psychol. 9: 343-347.
Deutsch, J. & Larsson, K. 1974. Model-oriented sexual behavior in surrogate-reared rhesus monkeys. Brain Behay. Evol. 9: 157-164.
Diner, J. 1979. Physical and mental suffering of experimental animals: A review of scientific literature 1975-1978. Washington, DC: Animal Welfare Institute.
Dolhinow, P. 1980. An experimental study of mother loss in the Indian langur monkey (Presbytis entellus). Folia Primatol. 33: 77-128.
Dolhinow, P. & Murphy, G. 1983. Langur monkey mother loss: profile analysis with multivariate analysis of variance for separation subjects and controls. Folia Primatol. 40: 181-196.
Drewett, R. & Kani, W. 1981. Animal experimentation in the behavioural sciences. Pp. 175-201 in: Animals in research (D. Sperlinger, ed.). New York: John Wiley & Sons, Ltd.
Eastman, R. F. & Mason, W. A. 1975. Looking behavior in monkeys raised with mobile and stationary artificial mothers. Dev. Psychobiol. 8: 213-221.
Elias, M. F. & Samonds, K. W. 1973. Exploratory behavior of cebus monkeys after having been reared in partial isolation. Child Devel. 44: 218-220.
Elliot, 0. & Scott, J. P. 1961. The development of emotional distress reactions to separation in puppies. J. Genet. Psychol. 99: 3-22.
Erwin, J. 1974. Responses of rhesus monkeys to separation vocalizations of a conspecific infant. Perc. Mot. Skills 39: 179-185.
Erwin, J., Maple, T., Mitchell, G. & Willott, J. 1974. Follow-up study of isolation-reared and mother-reared rhesus monkeys paired with preadolescent conspecifics in late infancy: Cross-sex pairings. Devel. Psychol. 10: 808-814.
Evans, C. S. 1967. Methods of rearing and social interaction in Macaca nemestrina. Anim. Behay. 15: 263-266.
Fittinghoff, N. A., Lindburg, D. G., Gomber, J. & Mitchell, G. 1974. Consistency and variability in the behavior of mature, isolation-reared, male rhesus macaques. Primates 15: 111-139.
Fittinghoff, N. A., Jr., Lindburg, D. G. & Mitchell, G. 1971. Failure to find polydipsia in isolation-reared monkeys. Psychonom. Sci. 22: 277-278.
Fitz-Gerald, F. L. 1966. Effects of d-amphetamine upon behavior of young chimpanzees reared under different conditions. Exc. Med. Int. Congr. Ser. 129: 1226-1227.
Floeter, M. K. & Greenough, W. T. 1979. Cerebellar plasticity: Modification of Purkinje cell structure by differential rearing in monkeys. Science 206: 227-228. Foley, J. P., Jr. 1934. First year development of a rhesus monkey (Macaca mulatta) reared in isolation. J. Genet. Psychol. 45: 39-105.
Foley, J. P., Jr. 1935. Second year development of a rhesus monkey (Macaca mulatta) reared in isolation during the first eighteen months. J. Genet. Psychol. 47: 73-97.
Fontana, V. J. 1973. Somewhere a child is crying. New York: MacMillan. Fox, M. W. 1966. A syndrome in the dog resembling human infantile autism. J. Am. Vet. Med. Assoc. 148: 1387-90.
Fox, M. W 1967. The effects of short-term social and sensory isolation upon behavior, EEG and averaged evoked potentials in puppies. Physiol. Behay. 2: 145-151.
Fox, M. W & Stelzner, D. 1966a. Spontaneous and experimentally induced behavioural abnormalities in the dog correlated with early experience and the critical period hypothesis. Pp. 29-49 in: Recent advances in biological psychiatry, Vol. 8. New York: Plenum.
Fox, M. W. & Stelzner, D. 1966b. Behavioural effects of differential early experience in the dog. Anim. Behay. 14: 273-281.
Frank, R. G., Gluck, J. P. & Strongin, T. S. 1977. Response suppression to a shock-predicting stimulus in differentially reared monkeys (Macaca mulatta). Devel. Psychol. 13: 295-296.
Frohman, C. E, Tourney, G., Beckett, P. G. S., Lees, H., Latham, L. K. & Gottlieb, J. S. 1961. Biochemical identification of schizophrenia. Arch. Gen. Psychiat. 4: 404-412.
Fuller, J. L. 1967. Experimental deprivation and later behavior. Science 158: 1645-52.
Fuller, J. L. & Clark, L. D. 1966a. Genetic and treatment factors modifying the post-isolation syndrome in dogs. J. Comp. Physiol. Psychol. 61: 251-257.
Fuller, J. L. & Clark, L. D. 1966b. Effects of rearing with specific stimuli upon post-isolation behavior in dogs. J. Comp. Psysiol. Psychol. 61: 258-263.
Fuller, J. L. & Clark, L. D. 1968. Genotype and behavioral vulnerability to isolation in dogs. J. Comp. Physiol. Psychol. 66: 151-156.
Furchner, C. S. & Harlow, H. F. 1969. Preference for various surrogate surfaces among infant rhesus monkeys. Psychonom. Sci. 17: 279-280.
Furman, R., Rahe, D. R. & Hartup, W. W. 1979. Rehabilitation of socially-withdrawn children through mixed-age and same-age socialization. Child Devel. 50: 915-922.
Gallup, G. G., Jr. & McClure, M. K. 1971. Preference for mirror-image stimulation in differentially reared rhesus monkeys. J. Comp. Physiol. 75: 403-407.
Gallup, G., McClure, M. K., Hill, S. D. & Bundy, R. A. 1971. Capacity for self-recognition in differentially reared chimpanzees. Psychol. Rec. 21: 69-74.
Gallup, G. & Suarez, S. 1980. On the use of animals in psychological research. Psychol. Rec. 30: 211-218.
Gendin, S. 1984. Letter. Agenda 6 (6): 3.
Gluck, J. P. 1979. The intellectual consequences of early social restriction in rhesus monkeys. Pp. 253-268 in: Nursery care of nonhuman primates (G. C. Ruppenthal & D. T. Reese, eds.) New York: Plenum Press.
Gluck, J. P. & Harlow, H. F. 1971. The effects of deprived and enriched rearing conditions on later learning: A review. Pp. 103-119 in: Cognitive processes of nonhuman primates (L. E. Jarrard, ed.). New York: Academic Press.
Gluck, J. P., Harlow, H. F. & Schiltz, K. A. 1973. Differential effect of early enrichment and deprivation on learning in the rhesus monkey. Comp. Physiol. Psychol. 84: 598-604.
Gluck, J. P. & Sackett, G. P. 1974. Frustration and self-aggression in social isolate rhesus monkeys. J. Abnorm. Psychol. 83: 331-334.
Gluck, J. P. & Sackett, G. P. 1976. Extinction deficits in a socially isolated rhesus monkey (Macaca mulatta). Devel. Psychol. 12: 173-174.
Gomber, J. & Mitchell, G. 1974. Preliminary report on adult male isolated-reared rhesus monkeys caged with infants. Devel. Psychol. 10: 298.
Green, P. C. 1965. Influence of early experience and age on expression of affect in monkeys. J. Genet. Psychol. 106: 157-171.
Green, P. C. & Gordon, M. 1964. Maternal deprivation: Its influence on visual exploration in infant monkeys. Science 145: 292-294.
Griffin, G. A. & Harlow, H. F. 1966. Effects of three months of total social deprivation on social adjustment and learning in the rhesus monkey. Child Devel. 37: 533-547.
Gunner, M. R., Gonzalez, G. A., Goodlin, B. L. & Levine, S. 1981. Behavioral and pituitary-adrenal response during a separation period in infant rhesus macaques. Psychoneuroendocrin. 6: 66-75.
Guyot, G. W., Cross, H. A. & Bennett, T. L. 1980. Early social isolation of the domestic cat: Responses to separation from social and nonsocial rearing stimuli. Dev. Psychobiol. 13: 309-315.
Hamburg, D. A. 1970. Psychiatry as a behavioral science. New Jersey: Prentice-Hall, Inc.
Hansen, E. W. 1966. The development of maternal and infant behavior in the rhesus monkey. Behavior 27: 107-149.
Harlow, H. F. 1958. The nature of love. Am. Psychol. 13: 673-685.
Harlow, H. F. 1959a. Love in infant monkeys. Sci. Am. 200 (June): 68-74.
Harlow, H. F. 1959b. Basic social capacities of primates. Hum. Biol. 31: 40-53.
Harlow, H. F. 1960. Primary affectional patterns in primates. Am. J. Orthopsychiat. 30: 676-684.
Harlow, H. F. 1962a. Development of affectional patterns in infant monkeys. Pp. 75-88 in: The determinants of infant behavior, vol. II (B. M. Foss, ed.). New York: Wiley.
Harlow, H. F. 1962b. Development of affection in primates. Pp. 157-166 in: Roots of behavior (E. L. Bliss, ed.). New York: Harper.
Harlow, H. F. 1963. The maternal affectional system. Pp. 3-33 in: The determinants of infant behavior (B. M. Foss, ed.). New York: Wiley.
Harlow, H. F. 1964. Early social deprivation and later behavior in the monkey. Pp. 154-173 in: Unfinished tasks in the behavioral sciences (A. Abrams, H. H. Gurner & J. E. P. Tomal, eds.). Baltimore: Williams & Wilkins.
Harlow, H. F. 1965. Total social isolation: Effects on macaque behavior. Science 148: 66 (abstract).
Harlow, H. F. 1969. Age-mate or peer affectional system. Pp. 333-383 in: Advances in the study of behavior, Vol. 2 (D. S. Lehrman, R. A. Hinde & E. Shaw, eds.). New York: Academic.
Harlow, H. F. 1974. Induction and alleviation of depressive states in monkeys. Pp. 197-208 in: Ethology and psychiatry (N. F. White, ed.). Toronto, Canada: University of Toronto Press.
Harlow, H. F. 1976. Syndromes resulting from maternal deprivation: 1. Maternal and peer affectional deprivation in primates. Pp. 85-98 in: Experimental behavior: A basis for the study of mental disturbance (J. H. Cullen, ed.). New York: J. Wiley & Sons.
Harlow, H. F., Dodsworth, R. 0. & Harlow, M. K. 1965. Total social isolation in monkeys. Proc. Nat. Acad. Sci. 54: 90-97.
Harlow, H., Gluck, J. P. & Suomi, S. J. 1972. Generalization of behavioral data between nonhuman and human animals. Am. Psychol. 27: 709-716.
Harlow, H. F. & Griffin, G. A. 1965. Induced mental and social deficits in rhesus monkeys. Pp. 87-106 in: The biosocial basis of mental retardation (S. F. Osier & R. E. Cooke, eds.). Baltimore: Johns Hopkins.
Harlow, H. F. & Harlow, M. K. 1962a. The effect of rearing conditions on behavior. Bull. Menninger Clinic 26: 213-224.
Harlow, H. F. & Harlow, M. K. 1962b. Social deprivation in monkeys. Sci. Am. 207: 136-146.
Harlow, H. F. & Harlow, M. K. 1965. The affectional systems. Pp. 287-334 in: Behavior of nonhuman primates, Vol. 2 (A. M. Schrier, H. F. Harlow & F. Stollnitz, eds.). New York: Academic Press.
Harlow, H. F. & Harlow, M. 1966. Learning to love. Am. Scient. 54: 244-272.
Harlow, H. F. & Harlow, M. K. 1969. Effects of various mother-infant relationships on rhesus monkey behaviors. Pp. 15-36 in: Determinants of infant behavior, Vol. 4 (B. M. Foss, ed.). London: Methuen.
Harlow, H. F. & Harlow, M. K. 1971. Psychopathology in monkeys. Pp. 203-229 in: Experimental psychopathology (H. D. Kimmel, ed.). New York: Academic Press.
Harlow, H. F., Harlow, M. K., Dodsworth, R. D. & Arling, G. L. 1966. Maternal behavior of rhesus monkeys deprived of mothering and peer association. Proc. Am. Phil. Soc. 110: 58-66.
Harlow, H. R, Harlow, M. K. & Hansen, E. W. 1963. The maternal affectional system of rhesus monkeys. Pp. 254-281 in: Maternal behavior in mammals (H. L. Rheingold, ed.). New York: Wiley.
Harlow, H. F., Harlow, M. K., Schiltz, K. A. & Mohr, D. J. 1971a. The effects of early adverse and enriched environments on the learning ability of rhesus monkeys. Pp. 121-148 in: Cognitive processes of nonhuman primates (L. E. Jarrard, ed.). New York: Academic Press.
Harlow, H. F., Harlow, M. K. & Suomi, S. J. 1971b. From thought to therapy: lessons from a primate laboratory. Am. Scient. 59: 538-549.
Harlow, H. F. & McKinney, W T. 1971. Nonhuman primates and psychosis. J. Aut. Child Schizo. 1: 368-375.
Harlow, H. F. & Novack, M. H. 1973. Psychopathological perspective. Persp. Biol. Med. 16: 461-478.
Harlow, H. F., Plubell, P. E. & Baysinger, C. M. 1973. Induction of psychological death in rhesus monkeys. J. Aut. Child Schizo. 3: 299-307.
Harlow, H. F., Rowland, G. L. & Griffin, G. A. 1 964 . The effect of total social deprivation on the development of monkey behavior. Pp. 116-135 in: Recent research on schizophrenia (Psychiatric research report #19) (P. Solomon & B. C. Glueck, Jr., eds.). American Psychiatric Association.
Harlow, H., Schlitz, K. A. & Harlow, M. K. 1969. Effects of social isolation on the learning performance of rhesus monkeys. Pp. 178-185 in: Proceedings of the 2nd International Congress on Primatology, Vol. 1, Behavior (C. R. Carpenter, ed.). Basel: S. Karger.
Harlow, H. F. & Seay, B. 1966. Mothering in motherless-mother monkeys. Br. J. Soc. Psychiat. 1: 63-69.
Harlow, H. F. & Suomi, S. J. 1970. The nature of love-simplified. Am. Psychol. 25: 161-168.
Harlow, H. F. & Suomi, S. J. 1971a. Social recovery by isolation-reared monkeys. Proc. Nat. Acad. Sci. 68: 1534-1538.
Harlow, H. F. & Suomi, S. J. 1971b. Production of depression behaviors in young monkeys. J. Aut. Child Schiz. 1: 246-55.
Harlow, H. F. & Suomi, S. J. 1974. Induced depression in monkeys. Behay. Biol. 12: 273-296.
Harlow, H. F. & Suomi, S. J. 1976. Workshop 1: Instinctual and environmental learning. Pp. 61-74 in: Animal models in human psychobiology (G. Serban & A. Kling, eds.). New York: Plenum.
Harlow, H. F. & Zimmerman, R. R. 1958. The development of affectional responses in infant monkeys. Am. Phil. Soc. 3: 1017-1023.
Harlow, H. F. & Zimmerman, R. R. 1959. Affectional responses in the infant monkey. Science 130: 421-452. Harris, J. 1984. The child: Development from birth through adolescence. Englewood Cliffs, NJ: Prentice Hall.
Heath, R. G. 1972. Electroencephalographic studies in isolation-raised monkeys with behavioral impairment. Dis. Nerv. Sys. 33: 157-163.
Hebb, D. 0. 1958. The motivating effects of exteroceptive stimulation. Am. Psychol. 13: 109-113.
Helton, D. 1985. Mind over monkey: When a psychologist experiments on an animal, which reveals more about human behavior -the human or the animal? BBC Wildlife Mag., reprinted in Animals Agenda 5: 26-27.
Hennessy, M. B. 1984. Presence of companion moderates arousal of monkeys with restricted social experience. Physiol. Behay. 33: 693-698.
Hennessy, M. B., Kaplan, J. N., Mendoza, S. P., Lowe, E. S. & Levine, S. 1979. Separation distress and attachment in surrogate-reared squirrel monkeys. Physiol. Behay. 23: 1017-1023.
Herzog, M. & Hopf, S. 1983. Effects of species-specific vocalizations on the behavior of surrogate-reared squirrel monkeys. Behaviour 86: 197-214.
Hill, S. D., McCormack, S. A. & Mason, W. H. 1973. Effects of artificial mothers and visual experience on adrenal responsiveness of infant monkeys. Dev. Psychobiol. 6: 421-429.
Hinde, R. A. 1972. Mother-infant separation in rhesus monkeys. J. Psychosom. Res. 16: 227-228.
Hinde, R. A. 1974. Mother/infant relations in rhesus monkeys. Pp. 29-46 in: Ethology and psychiatry (N. F. White, ed.). Toronto, Canada: University of Toronto Press.
Hinde, R. A. 1976. The use of differences and similarities in comparative psychopathology. Pp. 187-202 in: Animal models in human psychobiology (G. Serban & A. Kling, eds.). New York: Plenum.
Hinde, R. A. 1977. Mother-infant separation and the nature of inter-individual relationships: Experiments with rhesus monkeys. Proc. Roy. Soc. B. 196: 29-50.
Hinde, R. A. & Davies, L. M. 1972a. Changes in mother-infant relationship after separation in rhesus monkeys. Nature 239: 41-42.
Hinde, R. A. & Davies, L. M. 1972b. Removing infant rhesus from mother for 13 days compared with removing mother from infant. J. Child Psychol. Psychiat. 138: 227-237.
Hinde, R. A., Leighton-Shapiro, M. E. & McGinnis, L. 1978. Effects of various types of separation experience on rhesus monkeys 5 months later. J. Child Psychol. Psychiat. 19: 199-211.
Hinde, R. A. & McGinnis, L. 1977. Some factors influencing the effects of temporary mother-infant separation: Some experiments with rhesus monkeys. Psychol. Med. 7: 197-212.
Hinde, R. A. & Spencer-Booth, Y. 1968. The study of mother-infant interaction in captive group-living rhesus monkeys. Proc. Roy. Soc. B 169: 177-201.
Hinde, R. A. & Spencer-Booth, Y. 1970. Individual differences in the responses of rhesus monkeys to a period of separation from their mothers. J. Child Psychol. Psychiat. 11: 159-176.
Hinde, R. A. & Spencer-Booth, Y. 1971. Effects of brief separation from mother on rhesus monkeys. Science 173: 111-118.
Hinde, R. A., Spencer-Booth, Y. & Bruce, M. 1966. Effects of 6-day maternal deprivation on rhesus monkey infants. Nature 210: 1021-1023.
Hofer, M. A. 1970. Physiological responses of infant rats to separation from their mothers. Science 168: 871-873.
Hofer, M. A. 1971. Regulation of cardiac rate by nutritional factor in young rats. Science 172: 1039-41.
Hofer, M. A. 1973a. Physiological and behavioural processes in early maternal deprivation. Pp. 175-200 in: Physiology, emotion and psychosomatic medicine. Amsterdam: Elsevier/Excerpt Media/North-Holland (Ciba Foundation Symposium 8).
Hofer, M. A. 1973b. The role of nutrition in the physiological and behavioral effects of maternal separation on two week old rat pups. Psychosom. Med. 35: 350-359. Hofer, M. A. 1973c. Maternal separation affects infant rats’ behavior. Behay. Biol. 9: 629-633.
Hofer, M. A. 1973d. The effects of brief maternal separations on behavior and heart rate of two week old rat pups. Physiol. Behay. 10: 423-427. Hofer, M. A. 1975a. Studies on how early maternal separation produces behavioral change in young rats. Psychosom. Med. 37: 245-264.
Hofer, M. A. 1975b. Survival and recovery of physiologic function after early maternal separation in young rats. Physiol. Behay. 15: 475-480.
Hofer, M. A. 1975c. infant separation responses and the maternal role. Biol. Psychiat. 10: 149-53.
Hofer, M. A. 1976a. Olfactory denervation: its biological and behavioral effects on infant rats. J. Comp. Physiol. Psychol. 90: 829-838.
Hofer, M. A. 1976b. The organization of sleep and wakefulness after maternal separation in young rats. Dev. Psychobiol. 9: 189-205.
Hofer, M. A. 1978. Hidden regulatory processes in early social relationships. Pp. 135-166 in: Perspectives in ethology: II. Social behavior (P. P. Bateson & P. H. Klopfer, eds.). New York: Plenum.
Hofer, M. A. & Shair, H. 1982. Control of sleep-wake states in the infant rat by features of the mother-infant relationship. Dev. Psychobiol. 15: 229-243.
Hofer, M. A. & Weiner, H. 1971. The development and mechanisms of cardiorespiratory responses to maternal deprivation in rat pups. Psychosom. Med. 33: 353-362.
Hofer, M. A. & Weiner, H. 1975. Physiological mechanisms for cardiac control by nutritional intake after early maternal separation in the young rat. Psychosom. Med. 37: 8-24.
Hopf, S., Herzog, M. & Vogl-Koehler, C. 1984. Social integration of surrogate-reared infant squirrel monkeys to captive groups, Acta Paedopsychiat. 50: 79-95.
Horenstein, D. 1977. The dynamics and treatment of child abuse: Can primate research provide the answers? J. Clin. Psychiat. 33: 563-565.
Hrdina, P. D., von Kulmiz, P. & Stretch, R. 1979. Pharmacological modification of experimental depression in infant macaques. Psychopharmacol. 64: 89-93.
Hubbard, R. C. 1985. Life, funds wasted in areas of research. Worcester Gazette (Mass.), April 4: 29.
Huebner, D. K. & King, J. E. 1984. Kittens as therapists: Social behavior sequences in isolated squirrel monkeys after exposure to young nonconspecifics. Dev. Psychobiol. 17: 233-242.
Igel, G. J. & Calvin, H. D. 1960. The development of affectional responses in infant dogs. J. Comp. Physiol. Psychol. 53: 302-305.
Ingmanson, E. J. 1984. Behavioral responses to maternal separation in captive chimpanzee infants. Am. J. Phys. Anthrop. 63: 173-174.
Jensen, G. D. 1968. Reaction of monkey mothers to long-term separation from their infants. Psychon. Sci. 11: 171-172.
Jensen, G. D. & Tolman, C. W. 1962a. Mother-infant relationship in the monkey, Macaca nemestrina: The effect of brief separation and mother-infant specificity. J. Comp. Physiol. Psychol. 55: 131-136.
Jensen, G. & Tolman, C. 1962b. Activity level of the mother monkey, Macaca nemestrina, as affected by various conditions of sensory access to the infant following separation. Anim. Behay. 10: 228-230.
Jones, B. C. & Clark, D. L. 1973. Mother-infant separation in squirrel monkeys living in a group. Dev. Psychobiol. 6: 259-269.
Jordan, T. C., Coe, C. L., Patterson, J. & Levine, S. 1984. Predictability and coping with separation in infant squirrel monkeys. Behay. Neurosci. 98: 556-560.
Kaplan, J. 1970. The effects of separation and reunion on the behavior of mother and infant squirrel monkeys. Dev. Psychobiol. 3: 43-52. Kaplan, J. 1974. Growth and behavior of surrogate-reared squirrel monkeys. Dev. Psychobiol. 7: 7-13.
Kaplan, J. N. 1981. Effects of surrogate-administered punishment on surrogate contact in infant squirrel monkeys. Dev. Psychobiol. 14: 523-532.
Karrer, R. & Dekker, L. 1973. Preliminary observations on the development of dominance in partial-isolation-reared monkeys (Macaca irus). Bull. Psychonom. Soc. 2: 225-228.
Kaufman, I. C. 1973. Mother-infant separation in monkeys: An experimental model. Pp. 33-52 in: Separation and depression: Clinical and research aspects (J. P. Scott & E. C. Senay, eds.). Washington, D. C.: American Association for the Advancement of Science, No. 94.
Kaufman, 1. C. 1974. Mother/infant relations in monkeys and humans: A reply to Professor Hinde. Pp. 47- 68 in: Ethology and psychiatry (N. F. White, ed.). Toronto, Canada: University of Toronto.
Kaufman, I. C. 1977. Developmental considerations of anxiety and depression: Psychobiological studies of monkeys. Pp. 317-363 in: Psychoanalysis and contemporary science, Vol. 5 (T. Shapiro, ed.). New York: International Universities.
Kaufman, I. C. & Rosenblum, L. A. 1967a. The reaction to separation in infant monkeys: Anaclitic depression and conservation-withdrawal. Psychosom. Med. 29: 648-675.
Kaufman, I. C. & Rosenblum, L. A. 1967b. Depression in infant monkeys separated from their mothers. Science 155: 1030-1031.
Kaufman, I. C. & Rosenblum, L. A. 1969. Effects of separation from mother on the emotional behavior of infant monkeys. Ann. N. Y. Acad. Sci. 159: 681-695.
Kaufman, I. C. & Stynes, A. J. 1978. Depression can be induced in a bonnet macaque infant. Psychosom. Med. 40: 71-75.
Kawabe, S. 1969. Development of vocalization in monkeys reared in isolation. Int. J. Am. Ling. 35: 260 (abs.).
Kerr, G. R., Chamove, H. S. & Harlow, H. F. 1969. Environmental deprivation: Its effects on the growth of infant monkeys. J. Pediat. 75: 833-837.
Koch, M. D. & Arnold, W. J. 1972. Effects of early social deprivation on emotionality in rats. J. Comp. Physiol. Psychol. 78: 391-399.
Koch, M. D. & Arnold, W J. 1976. Maternal and nutritional factors in maintenance of infant rat cardiac rate following maternal separation. Physiol. Behav. 16: 521-527.
Kolb, L. & Brodie, K. 1982. Modern clinical psychiatry. Philadelphia: W. B. Saunders Co.
Kornetsky, C. 1977. Future directions. Pp. 497-499 in: Animal models in psychiatry and neurology (I. Hanin & E. Usdin, eds.). Oxford: Pergamon.
Kraemer, A. W., Ebert, M. H. & McKinney, W. T. 1983. Separation models and depression. Pp. 133-145 in: The origins of depression: Current concepts and approaches. (T. Angst, ed.). New York: Springer-Verlag.
Kraemer, A. & McKinney, W. 1979. Interaction of pharmacological agents which alter biogenic amine metabolism and depression. J. Affect. Disorders 1: 33-54.
Krushinskii, L. V. 1962. Animal behavior: Its normal and abnormal developments. New York: Consultants Bureau.
Kuker-Reines, B. 1982. Psychology experiments on animals. Boston: New England Anti-Vivisection Society.
Laudenslager, M. L. & Reite, M. L. 1984. Losses and separations: Immunological consequences and health implications. Pp. 285-312 in: Review of personality and social psychology, no. 5: Emotions, relationships, and health (P. Shaver, ed.). Beverly Hills: Sage Publications.
Laudenslager, M. L., Reite, M. & Harbeck, R. J. 1982. Suppressed immune response in infant monkeys associated with maternal separation. Behav. Neur. Biol. 36: 40-48.
Lebovici, S. 1962. The concept of maternal deprivation: A review of research. Pp. 75-95 in: Deprivation of maternal care: A reassessment of its effects. Geneva: WHO Public Health Papers, No. 14
Lehrmann, D. S. 1974. Can psychiatrists use ethology? Pp. 187-196 in: Ethology and psychiatry (N. F. White, ed.). Toronto, Canada: University of Toronto.
Lessac, M. S. & Solomon, R. L. 1969. Effects of early isolation on the later adaptive behavior of beagles. Dev. Psychol. 1: 14-25.
Levine, R. 1985. A primer on the ethical underpinnings of institutional review boards. Workshop presented at Conference of Public Responsibility in Medicine and Research, March 25-26, Boston, MA.
Levine, S., Coe, C. L., Smotherman, W P. & Kaplan, J. N. 1978. Prolonged cortisol elevation in the infant squirrel monkey after reunion with mother. Physiol. Behav. 20: 7-10.
Levine, S., Franklin, D. & Gonzalez, C. A. 1984. Influence of social variables on the biobehavioral response to separation in rhesus monkey infants. Child Devel. 55: 1386-1393.
Levy, D. M. 1952. Animal psychology in its relation to psychiatry. Pp. 483-507 in: Dynamic psychiatry (F. Alexander, ed.). Chicago: University of Chicago.
Lewis, J. K. 1978. Differential response to maternal separation as a function of separation environment in the rhesus monkey. Diss. Abs. Int. 38: 4508B.
Lewis, J. K., McKinney, W. T., Young, L. D. & Kraemer, G. W. 1976. Mother-infant separation in rhesus monkeys as a model of human depression: A reconsideration. Arch. Gen. Psychiat. 33: 699-705.
Lichstein, L. & Sackett, G. P. 1971. Reactions by differentially raised rhesus monkeys to noxious stimulation. Dev. Psychobiol. 4: 339-352.
Maple, T., Erwin, J. & Mitchell, G. 1974. Sexually aroused self-aggression in a socialized adult male monkey. Arch. Sex. Behav. 3: 471-475.
Mason, W 1960. The effects of social restriction on the behavior of rhesus monkeys. I. Free social behavior. J. Comp. Physiol. Psychol. 53: 582-589.
Mason, W. 1961a. The effects of social restriction on the behavior of rhesus monkeys. II. Tests of gregariousness. J. Comp. Physiol. Psycho!. 54: 287-290.
Mason, W A. 1961b. The effects of social restriction on the behavior of rhesus monkeys. Ill. Dominance tests. J. Comp. Physiol. Psychol. 54: 694-699.
Mason, W. A. 1961c. Effects of age and stimulus characteristics on manipulatory responsiveness of monkeys raised in a restricted environment. J. Genet. Psychol. 99: 301-308.
Mason, W. A. 1963a. Social development of rhesus monkeys with restricted social experience. Perc. Mot. Skills. 16: 262-270.
Mason, W A. 1963b. The effects of environmental restriction on the social development of rhesus monkeys. Pp. 151-173 in: Primate social behavior (C. H. Southwick, ed.). Princeton, N. J.: Van Nostrand.
Mason, W A. 1967. Motivational aspects of social responsiveness in young chimpanzees. Pp. 103-126 in: Early behavior: Comparative and developmental approaches (W. Stevenson, E. H. Hess & H. L. Reingold, eds.). New York: Wiley.
Mason, W. A. 1968. Early social deprivation in the nonhuman primates: Implications for human behavior. Pp. 70-101 in: Environmental influences (D. C. Glass, ed.). New York: Rockefeller University and Russell Sage Foundation.
Mason, W. A. 1971. Motivational factors in psychosocial development. Pp. 35-67 in: Nebraska symposium on motivation, 1970 (W J. Arnold & M. M. Page, eds.). Lincoln: University of Nebraska Press.
Mason, W. A. 1973. Regulatory functions of arousal in primate psychosocial development. Pp. 19-33 in: Behavioral regulators of behavior in primates (C. R. Carpenter, ed.). Lewisburg, PA: Bucknell University Press.
Mason, W A. & Berkson, G. 1962. Conditions influencing vocal responsiveness of infant chimpanzees. Science 137: 127-128.
Mason, W A. & Berkson, G. 1975. Effects of maternal mobility on the development of rocking and other behaviors in rhesus monkeys: A study with artificial mothers. Dev. Psychobiol. 8: 197-211.
Mason, W. A., Davenport, R. K., Jr. & Menzel, E. W, Jr. 1968. Early experience and the social development of rhesus monkeys and chimpanzees. Pp. 440-480 in: Early experience and behavior (G. Newton & S. Levine, eds.). Springfield, C. Thomas.
Mason, W. A. & Fitz-Gerald, E L. 1962. Intellectual performance of an isolation-reared rhesus monkey. Perc. Mot. Skills 15: 594.
Mason, W A. & Green, R C. 1962. The effects of social restriction on the behavior of rhesus monkeys. IV: Responses to a novel environment and to an alien species. J. Comp. Physiol Psychol. 55: 363-368.
Mason, W. A., Hill, S. D. & Thomsen, C. E. 1971. Perceptual factors in the development of filial attachment. Pp. 125-133 in: Proceedings of the 3rd International Congress of Primatology, Zurich 1970, Vol. 3 (H. Kummer, ed.). Basel: S. Karger.
Mason, W A., Hill, S. D. & Thomsen, C. E. 1974. Perceptual aspects of filial attachments in monkeys. Pp. 84-93 in: Ethology and psychiatry (N. F. White, ed.). Toronto, Canada: Univ. of Toronto Press.
Mason, W A. & Kenney, M. D. 1974. Redirection of filial attachments in rhesus monkeys: Dogs as mother surrogates. Science 183: 1209-11.
Mason, W. A. & Sponholz, R. R. 1963. Behavior of rhesus monkeys raised in isolation. J. Psychiat. Res. 1: 299-306.
McArdle, J. 1984. Psychological experimentation on animals: Not necessary, not valid. Humane Soc. News, Spring: 1-3.
McCullock, T. L. & Haslerad, G. M. 1939. Affective responses of an infant chimpanzee reared in isolation from its kind. J. Comp. Psychol. 28: 437-445.
McGinnis, L. M. 1978. Maternal separations in rhesus monkeys within a social context. J. Child Psychol. Psychiat. 19: 313-327.
McGinnis, L. M. 1979. Maternal separation vs. removal from group companions in rhesus monkeys. J. Child Psychol. Psychiat. 20: 15-27.
McGinnis, L. M. 1980. Maternal separation studies in children and nonhuman primates. Pp. 311-335 in: Maternal influences and early behavior (R. W Bell & W P. Smotherman, eds.). New York: Spectrum.
McKinney, W. T., Jr. 1974a. Animal models in psychiatry. Persp. Biol. Med. 17: 529-541.
McKinney, W. T. 1974b. Primate social isolation: Psychiatric implications. Arch. Gen. Psychiat. 31: 422-426.
McKinney, W. T. 1977a. Biobehaviorial models of depression in monkeys. Pp. 117-126 in: Animal models in psychiatry (I. Hanin & E. Usdin, eds.). Oxford: Pergamon.
McKinney, W. T. 1977b. Animal behavioral biological models relevant to depressive and affective disorders in humans. Pp. 107-122 in: Depression in childhood: Diagnosis, treatment, and conceptual models (J. G. Schulterbrandt & A. Raskin, eds.). New York: Raven.
McKinney, W. T. & Bunney, W. E. 1969. Animal model of depression. I. Review of evidence: Implications for research. Arch. Gen. Psychiat. 21: 240-248.
McKinney, W. T., Eising, R. G., Moran, E. C., Suomi, S. J. & Harlow, H. F. 1971. Effects of reserpine on the social behavior of rhesus monkeys. Dis. Nerv. Syst. 32: 735-741.
McKinney, W. T., Suomi, S. J. & Harlow, H. F. 1971. Depression in primates. Amer. J. Psychiat. 127: 1313-1320.
McKinney, W T., Jr., Suomi, S. J. & Harlow, H. F. 1973c. New models of separation and depression in rhesus monkeys. Pp. 53-66 in: Separation and depression: Clinical and research aspects (J. P. Scott & E. C. Senay, eds.). Washington, DC: American Association for the Advancement of Science, Publication No. 94.
McKinney, W. T., Suomi, S. J. & Harlow, H. F. 1975. Experimental psychopathology in nonhuman primates. Pp. 310-334 in: American handbook of psychiatry, Vol. 6: New psychiatric frontiers, 2nd ed. (D. Hamburg & H. K. Brodie, eds.). New York: Basic Books.
McKinney, W. T., Suomi, S. J., Kliese, K. A. & Moran, E . C. 1973b. Can psychopathology be re-induced in rhesus monkeys? Arch. Gen. Psychiat. 29: 630-634.
McKinney, W T., Young, L. D., Davies, J. M. & Suomi, S. J. 1973a. Chloropromazine treatment of disturbed monkeys. Arch. Gen. Psychiat. 29: 490-494.
Meier, G. W 1965a. Other data on the effects of social isolation during rearing upon reproductive behavior in the rhesus monkey (M. mulatta). Anim. Behay. 13: 228-231.
Meier, G. W. 1965b. Maternal behavior of feral- and laboratory-reared monkeys following the surgical delivery of their infants. Nature 206: 492-493.
Meier, G. W. 1970. Comments on Sackett’s “Innate mechanisms, rearing conditions and a theory of early experience in primates.” Pp. 55-60 in: The prediction of behavior: Early experience variables (M. R. Jones, ed.). Coral Gables: University of Miami Press.
Melzack, R. 1954. The genesis of emotional behavior: An experimental study of the dog. J. Comp. Physiol. Psychol. 47: 166-168.
Melzack, R. 1962. Effects of early perceptual restriction on simple visual discrimination. Science 137: 978-979.
Melzack, R. 1965. Effects of early experience on behavior: Experimental and conceptual considerations. Pp. 271-299 in: Psychopathology of perception (P. H. Hock & J. Zubin, eds.). New York: Grune and Stratton.
Melzack, R. & Burns, S. K. 1963. Neuropsychological effects of early sensory restriction. Boln. Inst. Estud. Med. Biol., Mex. 21: 407-425.
Melzack, R. & Scott, T. H. 1957. The effect of early experience and response to pain. J. Comp. Physiol. Psychol. 50: 155-161.
Melzack, R. & Thompson, W. R. 1956. Effects of early experience on social behavior. Can. J. Psychol. 10: 82-90.
Mendoza, S. R et. al. 1978. Pituitary-adrenal response to separation in mother and infant squirrel monkeys. Dev. Psychobiol. 11: 169-175.
Mendoza, S. P., Coe, C. L., Smotherman, W. P., Kaplan, J. & Levine, S. 1980. Functional consequences of attachment: A comparison of two species. Pp. 235-252 in: Maternal influences and early behavior (R. W Bell & W. P. Smotherman, eds.). New York: Spectrum.
Menzel, E. W., Jr. 1963. The effects of cumulative experience on response to novel objects in young isolation-reared chimpanzees. Behaviour 21: 1-12.
Menzel, E. W., Jr. 1964. Patterns of responsiveness in chimpanzees reared through infancy under conditions of environmental restriction. Psychol. Forsch. 27: 337-365.
Menzel, E. W., Jr., Davenport, R. K., Jr. & Rogers, C. M. 1963a. The effects of environmental restriction upon the chimpanzee’s responsiveness to objects. J. Comp. Physiol. Psychol. 56: 78-85.
Menzel, E. W., Jr., Davenport, R. K., Jr. & Rogers, C. M. 1963b. Effects of environmental restriction upon the chimpanzee’s responsiveness in novel situations. J. Comp. Physiol. Psychol. 56: 329-334.
Menzel, E. W., Davenport, R. K., Jr. & Rogers, C. M. 1970. The development of tool-using in wild-born and restriction-reared chimpanzees. Folia Primatol. 12: 273-283.
Meyer, J. S. & Bowman, R. E. 1972. Rearing experience, stress, and adrenocorticosteroids in the rhesus monkey. Physiol. Behay. 8: 339-343.
Meyer, J. S., Novack, M. A., Bowman, R. E. & Harlow, H. F. 1975. Behavioral and hormonal effects of attachment object separation in surrogate-peer-reared and mother-reared infant rhesus monkeys. Dev. Psychobiol. 8: 425-435.
Miller, R. E., Caul, W. F. & Mirsky, I. A. 1967. Communication of affects between feral and socially isolated monkeys. J. Pers. Soc. Psychol. 7: 231-239.
Miller, R. E., Caul, W. R. & Mirsky, I. A. 1971. Patterns of eating and drinking in socially-isolated rhesus monkeys. Physiol. Behay. 7: 128-134.
Miller, R. E., Mirsky, I. A., Caul, W. F. & Sakata, T. 1969. Hyperphagia and polydipsia in socially isolated rhesus monkeys. Science 165: 1027-1028.
Minami, T. 1977. Behavioral changes in mother-infant relations by 6-months separation in one-year-old Japanese monkeys. Ann. Anim. Psychol. 27: 11-20.
Mineka, S. 1982. Depression and helplessness in primates. Pp. 197-242 in: Child nurturance: Vol. 3. Studies of development in nonhuman primates (H. E. Fitzgerald, J. A. Mullins & P. Gage, eds.). N.Y.: Plenum.
Mineka, S. & Suomi, S. J. 1978. Social separation in monkeys. Psychol. Bill. 85: 1376-1400.
Mineka, S., Suomi, S. J. & DeLizio, R. D. 1981. Multiple separation in adolescent monkeys: An opponent-process interpretation. J. Exp. Psychol.: Gen. 110: 56-85.
Missakian, E. A. 1969. Reproductive behavior of socially deprived male rhesus monkeys. J. Comp. Physiol. Psychol. 69: 403-407.
Missakian, E. A. 1972. Effects of adult social experience on patterns of reproductive activity of socially deprived male rhesus monkeys (Macaca multatta). J. Pers. Soc. Psychol. 21: 131-134.
Mitchell, G. 1968a. Persistent behavior pathology in rhesus monkeys following early social isolation. Folia Primatol. 8: 132-147.
Mitchell, G. D. 1968b. Attachment differences in male and female infant monkeys. Child Dev. 39: 611-620.
Mitchell, G. D. 1970. Abnormal behavior in primates. Pp. 185-249 in: Primate behavior, Vol. 1 (L. Rosenblum, ed.). N.Y.: Academic.
Mitchell, G. 1972. Looking behavior in the rhesus monkey. J. Phenom. Psychol. 3: 53-67.
Mitchell, G. 1974. Syndromes resulting from social isolation-2. Primates. Pp. 216-223 in: Experimental behaviour: A basis for the study of mental disturbance (J. H. Cullen, ed.). New York: J. Wiley & Sons.
Mitchell, G. D. & Clark, D. L. 1968. Long term effects of social isolation in normally adapted rhesus monkeys. J. Gen. Psychol. 113: 117-128.
Mitchell, G. D., Harlow, H. F., Griffin, G. A. & Moller, G. W. 1967. Repeated maternal separation in the monkey. Psychon. Sci. 8: 197-198.
Mitchell, G. D., Raymond, E. J., Ruppenthal, G. C. & Harlow, H. F. 1966. Long term effects of total social isolation upon behavior of rhesus monkeys. Psychol. Rep. 8: 567-580.
Moore, A. U. 1968. Effects of modified maternal care in the sheep and goat. Pp. 481-529 in: Early experience and behavior (G. Newton & S. Levine, eds.). Springfield, C. Thomas.
Morrison, H. L., Kraemer, G. W. & McKinney, W. T. 1976. Protest-despair response to peer separation on rhesus monkeys and its potentiation by alteration of catecholamine metabolism. Psychosom. Med. 38: 66 (abs.).
Mussen, P. H., Kagan, J., Conger, J. & Huston, A. 1984. Child development and personality. New York: Harper & Row.
Nadler, R. D. & Codger, M. A. 1983. Maternal separation and reunion of an infant orangutan. Primates 24: 67-76.
Nadler, R. D. & Green, S. 1975. Separation and reunion of a gorilla infant and mother. Int. Zoo Yrbk. 15: 198-201.
National Institutes of Health, 1984. Research Grants, Fiscal Year 1983 Funds. Bethesda, MD: NIH (Publi-cation No. 84-1042).
Newell, T. G. 1967. Effects of maternal deprivation on later behavior in two inbred strains of mice. Psychon Sci. 9: 119-120.
Newman, J. D. & Symmes, D. 1974. Vocal pathology in socially deprived monkeys. Dev. Psychobiol. 7: 351-358.
Noble, A. B., McKinney, W. T., Jr., Mohr, C. & Moran, E. 1976. Diazepam treatment of socially isolated monkeys. Am. J. Psychiat. 133: 1165-70.
Novak, M. A. 1979. Social recovery of monkeys isolated for the first year of life: II. Long-term assessment. Dev. Psychol. 15: 50-61.
Novak, M. A. & Harlow, H. R. 1975. Social recovery of monkeys isolated for the first year of life. 1. Rehabilitation and therapy. Dev. Psychol. 11: 453-465.
Pettijohn, T. F. 1979. Attachment and separation distress in the infant guinea pig. Dev. Psychobiol. 12: 73-81
Pettijohn, T. F., Wong, T. W, Ebert, P. D. & Scott, J. P. 1977. Alleviation of separation distress in 3 breeds of young dogs. Dev. Psychobiol. 10: 373-381.
Plaut, S. M. & Davis, J. M. 1972. Effects of mother-litter separation on survival, growth, and brain amino-acid levels. Physiol. Behay. 8: 43-51.
Plaut, S. M., Thal, A., Haynes, E. D. & Wagner, J. E. 1974. Maternal deprivation in the rat: Prevention of mortality by nonlactating adults. Psychosom. Med. 36: 311-320.
Porter, R. H. 1974. Syndromes resulting from object deprivation – I. Environmental restrictions on chickens. Pp. 33-42 in: Experimental behaviour: A basis for the study of mental disturbance (J. H. Cullen, ed.). New York: J. Wiley & Sons.
Pratt, C. L. & Sackett, G. P. 1967. Selection of social partners as a function of peer contact during rearing. Science 155: 1133-35.
Pratt, D. 1980. Alternatives to pain in experiments on animals. New York: Argus Archives.
Preston, D. G., Baker, R. P. & Seay, B. M. 1970. Mother-infant separation in patas monkeys. Dev. Psychol. 3: 298-306.
Randolph, M. C. & Mason, W. A. 1969. Effects of rearing conditions on distress vocalizations in chimpanzees. Folia Primatol. 10: 103-112.
Reite, M. 1977. Maternal separation in monkey infants: A model of depression. Pp. 127-140 in: Animal models in psychiatry and neurology (J. Hanin & E. Usdin, eds.). N. Y.: Pergamon.
Reite, M. 1979. Towards a pathophysiology of grief. Psychosom. Med. 41: 78 (abstract). Reite, M. L., Horbeck, R. & Hoffman, A. 1981. Altered cellular immune response following peer separation. Life Sci. 29: 1133-36.
Reite, M., Kaufman, 1. C., Pauley, J. D. & Stynes, A. J. 1974. Depression in infant monkeys: Physiological correlates. Psychosom. Med. 36: 363-367.
Reite, M., Seiler, C., Crowley, T. J., Hydinger-MacDonald, M. & Short, R. 1982. Circadian rhythm changes following maternal separation in monkeys. Chronobiol. 9: 1-11.
Reite, M., Seiler, C. & Short, R. 1978. Loss of your mother is more than loss of a mother. Am. J. Psychiat. 135: 370-371.
Reite, M. & Short, R. 1977. Nocturnal sleep in isolation reared monkeys: Evidence for environmental independence. Dev. Psychobiol. 10: 555-561.
Reite, M. & Short, R. A. 1978. Nocturnal sleep in separated monkey infants. Arch. Gen. Psychiat. 35: 1247-53.
Reite, M. & Short, R. A. 1983. Maternal separation studies: Rationale and methodological considerations. Pp. 219-254 in: Ethopharmacology, primate models of neuropsychiatric disorders (K. A. Miczek, ed.). N. Y.: A. R. Liss.
Reite, M., Short, R., Kaufman, K., Stynes, A. J. & Pauley, J. D. 1978. Heart rate and body temperature in separated monkey infants. Biol. Psychiat. 13: 91-105.
Reite, M., Short, R. A. & Seiler, C. 1978. Physiological correlates of maternal separation in surrogate-reared infants: A study in altered attachment bonds. Dev. Psychobiol. 11: 427-435.
Reite, M., Short, R., Seiler, C. & Pauley, J. D. 1981. Attachment, loss, and depression. J. Child Psychol. Psychiat. 22: 141-169.
Reite, M. & Snyder, D. S. 1982. Physiology of maternal separation in a bonnet macaque infant. Am. J. Primat. 2: 115-120.
Rheingold, H. L. & Eckerman, C. 0. 1971. Familiar social and nonsocial stimuli and the kitten’s response to a strange environment. Dev. Psychobiol. 4: 71-89.
Rogers, C. M. & Davenport, R. K. 1969. Effects of restricted rearing on sexual behavior of chimpanzees. Dev. Psychol. 1: 200-204.
Rogers, C. M. & Davenport, R. K. 1971. Intellectual performance of differentially reared chimpanzees: Oddity. Am. J. Ment. Defic. 75: 526-530.
Rose, R. M., Mason, J. W & Brady, J. V. 1969. Adrenal responses to maternal separation and adaptation in experimentally-raised rhesus monkeys (Macaca mulatta). Pp. 211-218 in: Proceedings of the 2nd International Congress of Primatology, Vol. 1 (C. R. Carpenter, ed.). Basel: S. Karger.
Rosenblatt, J. S. 1974. Some features of early behavioural development in kittens. Pp. 94-110 in: Ethology and psychiatry (N. F. White, ed.). Toronto, Canada: Univ. of Toronto.
Rosenblatt, J., Turkewitz, G. & Schneirla, T. C. 1963. Early socialization in the domestic cat as based on feeding and other relationships between female and young. Pp. 51-74 in: Determinants of infant behavior (B. Foss, ed.). New York: Wiley.
Rosenblum, L. A. 1971. Infant attachment in monkeys. Pp. 85-113 in: The origin of human social relations (R. Schoffer, ed.). New York: Academic.
Rosenblum, L. A. & Harlow, H. F. 1963. Approach-avoidance conflict in the mother-surrogate situation. Psycho]. Rep. 12: 83-85.
Rosenblum, L. A. & Kaufman, I. C. 1968. Variations in infant development and response to maternal loss in monkeys. Am. J. Orthopsychiat. 38: 418-426.
Rosenblum, L. A. & Plimpton, E. H. 1981. The infant’s effort to cope with separation. Pp. 225-257 in: The uncommon child (M. Lewis & L. A. Rosenblum, eds.). N. Y.: Plenum.
Rosenblum, L. A. & Smiley, J. 1984. Therapeutic effects of an imposed foraging task in disturbed monkeys. J. Child Psychol. Psychiat. 25: 485-497.
Rowan, A. N. 1984. Of mice, models, & men: A critical evaluation of animal research. Albany: State University of New York Press. Roy, M. A. 1981. Abnormal behaviors in nursery-reared squirrel monkeys (S. sciureus). Am. J. Primatol. 1: 35-42.
Roy, M. A., Wolf, R. H., Martin, L. N., Rangan, S. R. S. & Allen, W. P. 1978. Social and reproductive behaviors in surrogate-reared squirrel monkeys (Saimiri sciureus). Lab. Anim. Sci. 28: 417-421.
Ruesch, H. 1982. Naked empress or the great medical fraud. Zurich: Buchverlag CIVIS Publications.
Ruesch, H. 1983. Slaughter of the innocent. New York: CIVITAS.
Ruppenthal, G. C., Arling, G. L., Harlow, H. F., Sackett, G. P. & Suomi, S. J. 1976. A 10-year perspective of motherless-mother monkey behavior. J. Abn. Psychol. 85: 341-349.
Ryder, R. D. 1983. Victims of science: The use of animals in research. London: National Anti-Vivisection Society, Ltd.
Sackett, G. P. 1965a. Effects of rearing conditions upon the behavior of rhesus monkeys. Child Dev. 36: 855-868.
Sackett, G. P. 1965b. Manipulatory behavior in monkeys reared under different levels of early stimulus variation. Perc. Mot. Skills 20: 985-988.
Sackett, G. P. 1966. Monkeys reared in isolation with pictures as visual input: Evidence for an innate releasing mechanism. Science 154: 1468-1472.
Sackett, G. R 1967a. Some persistent effects of deficient rearing conditions in preadult social behavior of monkeys. J. Comp. Physiol. Psychol. 64: 363-365.
Sackett, G. P. 1967b. Some effects of social and sensory deprivation during rearing on behavioral devel-opment of monkeys. Revista Interam. Psicol. 1: 55-80.
Sackett, G. P. 1968a. Abnormal behavior in laboratory-reared rhesus monkeys. Pp. 293-331 in: Abnormal behavior in animals (M. W. Fox, ed.). Philadelphia: Saunders.
Sackett, G. P. 1968b. The persistence of abnormal behavior in monkeys following isolation rearing. Pp. 3-25 in: The role of learning in psychotherapy (R. Porter, ed.). London: Churchill.
Sackett, G. P. 1970a. Unlearned responses, differential rearing experiences, and the development of social attachments by rhesus monkeys. Pp. 111-140 in: Primate behavior, Vol. 1. (L. A. Rosenblum, ed.). N. Y.: Academic.
Sackett, G. R 1970b. Innate mechanisms, rearing conditions, and a theory of early experience effects in primates. Pp. 11-53 in: Miami symposium on the prediction of behavior: Effects of early experience (M. R. Jones, ed.). Miami: University of Miami Press.
Sackett, G. R 1972a. Exploratory behavior of rhesus monkeys as a function of rearing experiences and sex. Dev. Psychol. 6: 260-270.
Sackett, G. P. 1972b. Prospects for research on schizophrenia: Isolation-rearing in primates. Neurosci. Res. Prog. Bull. 10: 388-392.
Sackett, G. P. 1972c. Isolation rearing in monkeys: Diffuse and specific effects on later behavior. Pp. 61-110 in: Modeles animaux du comportement humain (R. Chauvin, ed.). Paris: Colloques Internationaux du C.N.R.S.
Sackett, G. R 1974. Syndromes resulting from object deprivation -2. Effects on development of rhesus monkeys. Pp. 43-169 in: Experimental behaviour: A basis for the study of mental disturbance (J. H. Cullen, ed.). New York: J. Wiley & Sons.
Sackett, G. P., Bowman, R. E., Meyer, J. S., Tripp, R. L. & Grady, S. S. 1973. Adrenocortical and behavioral reactions by differentially raised rhesus monkeys. Physiol. Psychol. 1: 209-212.
Sackett, G. P., Griffin, G. A., Pratt, C., Joslyn, W. D. & Ruppenthal, G. C. 1967. Mother-infant and adult female choice behavior in rhesus monkeys after various rearing experiences. J. Comp. Physiol. Psychol. 63: 376-381.
Sackett, G. R, Holm, R. A. & Ruppenthal, G. C. 1976b. Social isolation rearing: Species differences in behavior of macaque monkeys. Dev. Psychol. 10: 283-288.
Sackett, G. P., Holm, R. A., Ruppenthal, G. C. & Fahrenbruch, C. E. 1976a. The effects of total isolation rearing on the behavior of rhesus and pigtail macaques. Pp. 115-131 in: Environments as therapy for brain dysfunction (N. E. Walsch & W. Greenough, eds.). New York: Plenum.
Sackett, G. R, Porter, M. & Holmes, H. 1965. Choice behavior in rhesus monkeys: Effects of stimulation during the first month of life. Science 147: 304-306.
Sackett, G. P. & Ruppenthal, G. C. 1974. Some factors influencing the attraction of adult female macaque monkeys to neonates. Pp. 163-185 in: The effect of the infant on its caregiver (M. Lewis & L. A. Rosenblum, eds.). New York: Wiley-Interscience.
Sackett, G. P., Ruppenthal, G. C., Fahrenbruch, C. E., Holm, R. A. & Greenough, W. T. 1981. Social isolation effects in monkeys vary with genotype. Dev. Psychol. 17: 313-318.
Schlottmann, R. S. & Seay, B. 1972. Mother-infant separation in the java monkey (Macaca irus). J. Comp. Physiol. Psychol. 79: 334-304.
Scott, J. P. 1971. Attachment and separation in dog and man: Theoretical propositions. Pp. 227-246 in: The origins of human social relations (R. Schaffer, ed.). New York: Academic.
Scott, J. P. & Bronson, F. H. 1964. Experimental exploration of the et-epimeletic or care-soliciting behavioral system. Pp. 174-193 in: Psychobiological approaches to social behavior (P. H. Leiderman & L. D. Shapiro, eds.). Stanford: Stanford Univ.
Scott, J. P., Stewart, J. M. & DeGhett, V. J. 1973. Separation in infant dogs: Emotional response and motivational consequences. Pp. 3-32 in: Separation and depression: clinical and research aspects (J. P. Scott & E. C. Senay, eds.). Washington, D. C.: American Association for the Advancement of Science, No. 94.
Seay, B., Alexander, B. K. & Harlow, H. F. 1964. The maternal behavior of socially deprived rhesus monkeys. J. Abnorm. Soc. Psychol. 69: 345-354.
Seay, B., Hansen, E. & Harlow, H. F. 1962. Mother-infant separation in monkeys. J. Child Psychol. Psychiat. 3: 123-132.
Seay, B. & Harlow, H. F. 1965. Maternal deprivation in the rhesus monkey. J. Nerv. Ment. Dis. 140: 434-441.
Seiler, C., Cullen, J. S., Zimmerman, J. & Reite, M. 1979. Cardiac arrhythmias in infant pigtail monkeys following maternal separation. Psychophysiol. 16: 130-135.
Seitz, R F. D. 1958. Infantile experiences and adult behavior in animal subjects: II. Age of separation from the mother and adult behavior in the cat. Psychosom. Med. 20: 410 (Abstract)
Seitz, P. F. D. 1959. Infantile experience and adult behavior in animal subjects: II. Age of separation from the mother and adult behavior in the cat. Psychosom. Med. 21: 353-378.
Senay, E. C. 1966. Toward an animal model of depression: A study of separation behavior in dog. J. Psychiat. Res. 4: 65-71.
Serban, G. 1976. The significance of ethology for psychiatry. Pp. 279-289 in: Animal models in human psychobiology (G. Serban & A. Kling, eds.). New York: Plenum.
Serban, G., Pichot, P., Freedman, A. F. & Kittay, S. 1976. New perspectives in psychiatry: Relevance of the psychopathological animal model to the human. Pp. 1-6 in: Animal models in human psychobiology (G. Serban & A. Kling, eds.). New York: Plenum.
Shaw, E. 1962. Environmental conditions in the appearance of sexual behavior in the platyfish. Pp. 123-141 in: Roots of behavior (E. L. Bliss, ed.). New York: Harper & Brothers.
Short, R., Iwata, S. & Reite, M. 1977. EEG changes during the depressive reaction following maternal separation. Psychophysiol. 14: 120 (abstract).
Simons, R. C., Bobbitt, R. A. & Jensen, G. D. 1968. Mother monkey (Macaca nemestrina) responses to infant vocalizations. Perc. Mot. Skills 27: 3-10.
Singh, M. 1975. Mother-infant separation in rhesus monkeys living in natural environments. Primates 16: 471-476.
Singh, S. D. 1977. Effects of infant-infant separation of young monkeys in a free-ranging natural environment. Primates 18: 205-214.
Smotherman, W R, Hunt, L. F., McGinnis, L. M. & Levine, S. 1979. Mother-infant separation in group-living rhesus macaques: A hormonal analysis. Dev. Psychobiol. 12: 211-217.
Snyder, D. S., Graham, C. E., Bowen, J. A. & Reite, M. 1984. Peer separation in infant chimpanzees, a pilot study. Primates 25: 78-88.
Spencer-Booth, Y. & Hinde, R. A. 1967. The effects of separating rhesus monkey infants from their mothers for six days. J. Child Psychol. Psychiat. 7: 179-197.
Spencer-Booth, Y. & Hinde, R. A. 1971a. Effects of six-days separation from mother on 18-32-week old rhesus monkeys. Anim. Behay. 19: 174 – 191 .
Spencer-Booth, Y. & Hinde, R. A. 1971b. The effects of 13 days maternal separation on 30-32 week-old rhesus monkeys compared with those of shorter and interrupted separations. Anim. Behay. 19: 595-605.
Spencer-Booth, Y. and Hinde, R. 1971c. Effects of brief separations from mothers during infancy on behavior of rhesus monkeys 6-24 months later. J. Child Psychol. Psychiat. 12: 157-172.
Spitz, R. A. & Wolf, K. M. 1946. Anaclitic depression. Psychoanal. Stud. Child. 2: 313-342.
Spodek, B. 1982. Handbook of research in early childhood education. New York: Free Press.
Steele, B. F. & Pollock, C. B. 1968. A psychiatric study of parents who abuse infants and small children. Pp. 103-147 in: The battered child (R. Halfer & C. Kempe, eds.). Chicago: University of Chicago Press.
Stone, E. R., Bonnett, K. H. & Hofer, M. A. 1976. Survival and development of maternally deprived rats: Role of body temperature. Psychosom. Med. 38: 242-249.
Suomi, S. J. 1973a. Surrogate rehabilitation of monkeys reared in total social isolation. J. Child Psychol. Psychiat. 14: 71-77.
Suomi, S. J. 1973b. Repetitive peer separation of young monkeys: effects of vertical chamber confinement during separations. J. Abn. Psychol. 81: 1-10.
Suomi, S. J. 1976a. Mechanisms underlying social development: A reexamination of mother-infant interactions in monkeys. Pp. 201-228 in: Minnesota symposium on child psychology, Vol. 10 (A. Pick, ed.). Minneapolis: University of Minnesota Press.
Suomi, S. J. 1976b. Factors affecting responses to social separation in rhesus monkeys. Pp. 9-26 in: Animal models in human psychobiology (G. Serban & A. Kling, eds.). New York: Plenum.
Suomi, S. J. 1977a. Neglect and abuse of infants by rhesus monkey mothers. Voices [J. of the American Academy of Psychotherapists] 12: 5-8.
Suomi, S. J. 1977b. The development of attachment and other social behaviors in rhesus monkeys. Pp. 197-224 in: Attachment behavior (L. Krames, T. Alloway & P. Pliner, eds.). New York: Plenum Press.
Suomi, S. J. 1978. Maternal behavior by socially incompetent monkeys: Neglect and abuse of offspring. J. Ped. Psychol. 3: 28-34.
Suomi, S. J. 1979. Peers, play and primary prevention primates. Pp. 127-149 in: Primary prevention of psychopathology, Vol. 3: Social competence in children (M. W. Kent & J. E. Rolf, eds.). Hanover, N. H.: Univ. Press of New England.
Suomi, S. J. 1981. Genetic, maternal, and environmental influences on social development in rhesus monkeys. Pp. 81-87 in: Primate behavior and sociobiology (B. Chiarelli & R. Corruccini, eds.). Berlin: Springer-Verlag.
Suomi, S. J. 1982. Relevance of animal models for clinical psychology. Pp. 249-271 in: Handbook of research in clinical psychology (P. C. Kendall & J. N. Butcher, eds.). New York: Wiley.
Suomi, S. J. 1983. Social development in rhesus monkeys: Consideration of individual differences. Pp. 71-92 in: The behaviour of human infants (A. Oliverio & M. Zappella, eds.). New York: Plenum Press.
Suomi, S. J. 1984a. Ethology: Animals models. Pp. 226-236 in: Comprehensive textbook of psychiatry (H. I. Kaplan & B. J. Sadock, eds.). Baltimore: William & Wilkins.
Suomi, S. J. 1984b. The development of affect in rhesus monkeys. Pp. 119-159 in: The psychobiology of affective development (N. A. Fox & R. J. Davidson, eds.). Hillsdale, NJ: L. Erlbaum Assoc.
Suomi, S. J. 1986. Anxiety-like disorders in young nonhuman primates. In: Anxiety disorders of childhood (R. Gittelman, ed.) New York: Guilford Press.
Suomi, S. J., Collins, M. L. & Harlow, H. F. 1973. Effects of permanent separation from mother on infant monkeys. Dev. Psychol. 9: 376-384.
Suomi, S. J., Collins, M. L. & Harlow, H. E 1976. Effect of maternal and peer separations on young monkeys. J. Child Psychol. Psychiat. 17: 101-112.
Suomi, S. J. & Harlow, H. F. 1969. Apparatus conceptualization for psychopathological research in monkeys. Behay. Res. Meth. Instr. 1: 1-12.
Suomi, S. J. & Harlow, H. F. 1971. Monkeys at play. Nat. Hist. 80: 72-76.
Suomi, S. J. & Harlow, H. F. 1972a. Social rehabilitation of isolate-reared monkeys. Dev. Psycho!. 6: 487-496.
Suomi, S. J. & Harlow, H. F. 1972b. Depressive behavior in young monkeys subjected to vertical chamber confinement. J. Comp. Physiol. Psycho’. 80: 11-18.
Suomi, S. J. & Harlow, H. F. 1975a. The role and reason of peer relationships in rhesus monkeys. Pp. 153-185 in: Friendship and peer relations (M. Lewis & L. A. Rosenblum, eds.). New York: J. Wiley & Sons.
Suomi, S. J. & Harlow, H. F. 1975b. Effects of differential removal from group on social development of rhesus monkeys. J. Child Psycho’. Psychiat. 16: 149-164.
Suomi, S. J. & Harlow, H. F. 1976. The facts and functions of fear. Pp. 3-34 in: Emotions and anxiety (M. Zuckerman & C. D. Speilberger, eds.). Hillsdale, NJ: L. Erlbaum Assoc.
Suomi, S. J. & Harlow, H. F. 1977a. Early separation and behavioral maturation. Pp. 197-214 in: Genetics, environment and intelligence (A. Oliverio, ed.). Amsterdam: Elsevier/North Holland Biomedical Press.
Suomi, S. J. & Harlow, H. F. 1977b. Production and alleviation of depressive behaviors in monkeys. Pp. 131-173 in: Psychopathology: Experimental models (J. D. Maser & M. E. P. Seligman, eds.). San Francisco: Freeman.
Suomi, S. J. & Harlow, H. F. 1978. Early experience and social development in rhesus monkeys. Pp. 252-271 in: Social and personality development (M. Lamb, ed.). N. Y.: Holt, Rinehart.
Suomi, S. J., Harlow, H. F. & Domek, C. J. 1970b. Effect of repetitive infant-infant separation of young monkeys. J. Abn. Psychol. 76: 161-172.
Suomi, S. J., Harlow, H. F. & Kimball, S. D. 1971. Behavioral effects of prolonged partial social isolation in the rhesus monkey. Psychol. Rep. 29: 1171-1177.
Suomi, S. J., Harlow, H. F. & McKinney, W. T. 1972. Monkey psychiatrists. Am. J. Psychiat. 128: 927-932.
Suomi, S. J., Harlow, H. F. & Novak, M. A. 1974. Reversal of social deficits produced by isolation-rearing in monkeys. J. Human Evol. 3: 527-534.
Suomi, S. J. & Immelmann, K. 1983. On the process and product of cross-species generalization. Pp. 203-224 in: Studying man studying animals (D. W. Rajecki, ed.). Hillsdale, NJ: L. Erlbaum Assoc.
Suomi, S. J., Kraemer, G. W, Baysinger, C. M. & Delizio, R. D. 1981. Inherited and experiential factors associated with individual differences in anxious behavior displayed by rhesus monkeys. Pp. 179-200 in: Anxiety: New research and changing concepts (D. Klein & J. Rabkin, eds.). New York: Raven Press.
Suomi, S. J., Mineka, S. & DeLizio, R. D. 1983. Short- and long-term effects of repetitive mother-infant separations on social development in rhesus monkeys. Dev. Psychol. 19: 770-786.
Suomi, S. J., Mineka S. & Harlow, H. F. 1983. Social separation in monkeys as viewed from several motivational perspectives. Pp. 543-583 in: Handbook of behavioral neurobiology: Motivation (P. Teitel-baum & E. Satinoff, eds.). New York: Plenum.
Suomi, S. J. & Ripp, C. 1983. A history of motherless mother monkeys mothering at the University of Wisconsin Primate Laboratory. Pp. 49-78 in: Child abuse: The nonhuman primate data (M. Reite & N. Caine, eds.). New York: Liss.
Suomi, S. J., Sackett, G. P. & Harlow, H. F. 1970a. Development of sex preferences in rhesus monkeys. Develop. Psychol. 3: 326-336.
Suomi, S. J., Seaman, S. F., Lewis, J. K., Delizio, R. B. & McKinney, W. T. 1978. Effects of imipramine treatment of separation-induced social disorders in rhesus monkeys. Arch. Gen. Psychiat. 35: 321-325.
Symmes, D., Eisengart, M. A. & Healy, M. E. 1971. Neurobehavioral studies of isolation-reared monkeys. Proc. Am. Psychol. Assoc. 6: 789.
Tavris, C. 1973. Harry, you are going to go down in history as the father of the cloth mother. Psychol. Today 6: 65-77.
Thompson, W. R. 1960. Early environmental influences on behavioral development. Am. J. Orthopsychiat. 30: 306-314.
Thompson, W. R. 1962. The effects of prenatal and early postnatal experience. Pp. 10-17 in: Lessons from animal behaviour for the clinician (S. A. Barnett, ed.). London: Heinemann.
Thompson, W R. & Heron, W. 1954. The effects of restricting early experience on the problem-solving capacity of dogs. Can. J. Psychol. 8: 17-31.
Thompson, W. R., Melzack, R. & Scott, T. H. 1956. “Whirling behavior” in dogs as related to early experience. Science 123: 939.
Torrey, L. 1984. The agony of primate research. Sci. Dig. 92: 70-72.
Tuma, J. 1982. Handbook for the practice of pediatric psychology. New York: Wiley.
Turner, C. H., Davenport, R. K. & Rogers, C. J. 1969. The effect of early deprivation on the social behavior of adolescent chimpanzees. Am. J. Psychiat. 125: 85-90.
Vogt, J. L., Coe, C. L., Lowe, E. & Levine, S. 1980. Behavioral and pituitary-adrenal response of adult squirrel mopnkeys to mother-infant separation. Psychoneuroendocrinol. 3: 181-190.
Vogt, J. L. & Levine, S. 1980. Response of mother and infant squirrel monkeys to separation and disturbance. Physiol. Behay. 24: 829-832.
White, N. F. 1974. Ethology and psychiatry. Pp. 3-25 in: Ethology and psychiatry (N. F. White, ed.). Toronto, Canada: Univ. Toronto Press.
Whitten, C. F., Fischoff, J., Pettit, M. & Chodorkoff, J. 1968. Evidence that growth retardation in the maternal deprivation syndrome is secondary to an inadequate intake of calories (undereating). J. Pediat. 72: 563-565.
Willner, P. 1984. The validity of animal models of depression. Psychopharmacol. 83: 1-16.
Wood, B. S., Mason, W. A. & Kenny, M. D. 1979. Contrasts in visual responsiveness and emotional arousal between rhesus monkeys raised with living and those raised with inanimate substitute mothers. J. Comp. Physiol. Psychol. 93: 368-377.
World Health Organization. 1962. Preface. Pp. 7-8 in: Deprivation of maternal care. A reassessment of its effects (Anonymous editor). Geneva: WHO Public Health Papers, No. 14.
Yarrow, L. J. 1961. Maternal deprivation: Toward an empirical and conceptual reevaluation. Psychol. Bull. 58: 459-490.
Yarrow, L. J. 1964. Separation from parents during early childhood. Pp. 89-136 in: Review of child development research, Vol. 1 (M. L. Hoffman & L. W. Hoffman, eds.). New York: Russell Sage Foundation.
Yarrow, L. J. 1968. The crucial nature of early experience. Pp. 101-113 in: Environmental influences (D. C. Glass, ed.). N. Y.: Rockefeller University Press and Russell Sage Foundation.
Young, L., McKinney, W. T., Lewis, J. L. et al. 1973. Induction of adrenal catecholamine synthesizing enzymes following mother-infant separation. Nature 246: 94-96.
Young, L. D., Suomi, S. J., Harlow, H. F. & McKinney, W. T. 1973. Early stress and later response to separation. Am. J. Psychiat. 130: 400-405.